
植物生态学报 ›› 2018, Vol. 42 ›› Issue (5): 539-549.DOI: 10.17521/cjpe.2018.0042
所属专题: 全球变化与生态系统
宋小艳1,2,王根绪1,*( ),冉飞1,杨燕1,张莉1,2,肖瑶1,2
),冉飞1,杨燕1,张莉1,2,肖瑶1,2
收稿日期:2018-02-13
修回日期:2018-05-09
出版日期:2018-05-20
发布日期:2018-07-20
通讯作者:
王根绪
基金资助:
SONG Xiao-Yan1,2,WANG Gen-Xu1,*(),RAN Fei1,YANG Yan1,ZHANG Li1,2,XIAO Yao1,2
Received:2018-02-13
Revised:2018-05-09
Online:2018-05-20
Published:2018-07-20
Contact:
Gen-Xu WANG
Supported by:摘要:
为了揭示暖干化气候变化趋势对演替初期泰加林灌草层植物开花物候和生长的影响, 以东北大兴安岭演替初期泰加林灌草层(高度<50 cm)植物为研究对象, 根据植物生活史, 选择典型的早花植物圆锥薹草(Carex diandra)和水葡萄茶藨子(Ribes procumbens), 中花植物杜香(Ledum palustre)及晚花植物齿叶风毛菊(Saussurea neoserrata), 监测模拟增温和排水处理2年后其完整的开花物候及生长状况, 以期为认识气候变化下泰加林火后植被恢复演替提供依据。结果显示: 1)模拟增温和排水处理后, 早花植物开花温度限制的提早解除使其花期提前, 而水分限制使得晚花植物花期呈延后趋势, 这可能为中花植物提供更大的生态位空隙, 使其开花数量趋向于增加, 花期提前且呈延长趋势; 2)模拟增温和排水处理后, 圆锥薹草、杜香和齿叶风毛菊的盖度和频度均呈增加的趋势, 以杜香最显著, 而水葡萄茶藨子的盖度和频度均下降; 3)模拟增温与排水处理对植物开花物候的影响无显著交互作用, 但土壤水分降低影响部分植物生长对增温的响应。研究结果表明, 植物开花物候对暖干化气候的响应表现出明显的种间差异, 因植物开花功能群而异。早、中花植物花期趋于提前, 晚花期植物花期趋于延后, 一方面可能导致群落生态位变化, 对种间竞争产生潜在影响, 引发群落组成和结构改变; 另一方面可能成为群落调节其整体物候, 以适应气候变化的重要途径。
宋小艳, 王根绪, 冉飞, 杨燕, 张莉, 肖瑶. 东北大兴安岭演替初期泰加林灌草层典型植物开花物候与生长对模拟暖干化气候的响应. 植物生态学报, 2018, 42(5): 539-549. DOI: 10.17521/cjpe.2018.0042
SONG Xiao-Yan, WANG Gen-Xu, RAN Fei, YANG Yan, ZHANG Li, XIAO Yao. Flowering phenology and growth of typical shrub grass plants in response to simulated warmer and drier climate in early succession Taiga forests in the Da Hinggan Ling of northeast China. Chinese Journal of Plant Ecology, 2018, 42(5): 539-549. DOI: 10.17521/cjpe.2018.0042
| 对照 Control | 增温 Warming | 排水 Drainage | 增温+排水 Warming + Drainage | |
|---|---|---|---|---|
| 空气温度 Air temperature (℃) | 7.62 | 8.36 | 7.69 | 8.68 |
| 土壤温度 Soil temperature at 5 cm (℃) W, D | 5.37 ± 0.14 | 5.38 ± 0.14 | 5.97 ± 0.04 | 5.91 ± 0.13 |
| 土壤水分 Soil moisture at 0-10 cm (%)W, D | 45.52 ± 2.31 | 38.16 ± 6.82 | 25.36 ± 2.47 | 19.52 ± 1.05 |
| 最大融化深度 Maximun thaw depth (cm)D | 62.4 ± 1.2 | 65.1 ± 1.2 | 57.5 ± 1.8 | 55.2 ± 1.6 |
| 生长季平均水位 Mean water table (cm) | -20.0 | ND | -26.3 | ND |
表1 模拟增温和排水处理对生长季主要环境因子的影响(平均值或平均值±标准误差)
Table 1 Simulative warming and drainage effects on the most important environmental factors (mean or mean ± SE)
| 对照 Control | 增温 Warming | 排水 Drainage | 增温+排水 Warming + Drainage | |
|---|---|---|---|---|
| 空气温度 Air temperature (℃) | 7.62 | 8.36 | 7.69 | 8.68 |
| 土壤温度 Soil temperature at 5 cm (℃) W, D | 5.37 ± 0.14 | 5.38 ± 0.14 | 5.97 ± 0.04 | 5.91 ± 0.13 |
| 土壤水分 Soil moisture at 0-10 cm (%)W, D | 45.52 ± 2.31 | 38.16 ± 6.82 | 25.36 ± 2.47 | 19.52 ± 1.05 |
| 最大融化深度 Maximun thaw depth (cm)D | 62.4 ± 1.2 | 65.1 ± 1.2 | 57.5 ± 1.8 | 55.2 ± 1.6 |
| 生长季平均水位 Mean water table (cm) | -20.0 | ND | -26.3 | ND |
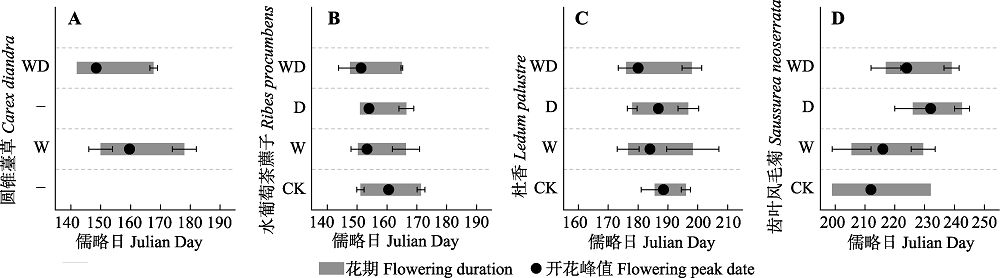
图1 模拟增温和排水处理对灌草层代表性物种开花物候的影响(平均值±标准误差)。CK、W、D和WD分别代表对照、增温、排水和增温+排水处理。
Fig. 1 Simulated warming and drainage effects on the flowering phenology of the typical shrub-grass species (mean ± SE). CK, control; W, warming; D, drainage; WD, warming + drainage.
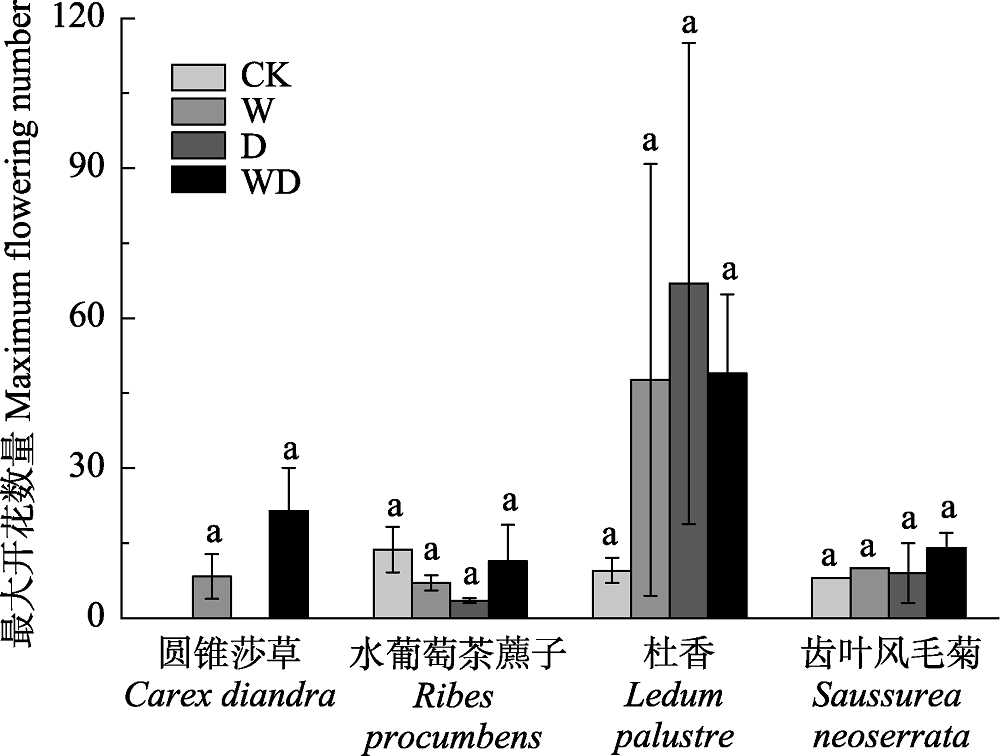
图2 模拟增温和排水处理对灌草层代表性物种最大开花数量的影响(平均值±标准误差)。相同物种内不同小写字母表示处理间差异显著(p < 0.05)。CK、W、D和WD分别代表对照、增温、排水和增温+排水处理。
Fig. 2 Simulated warming and drainage effects on maximum flowering number of typical shrub grass species (mean ± SE). Different small letters in the same species indicated significant difference among different treatments (p < 0.05). CK, control; W, warming; D, drainage; WD, warming + drainage.
| 物种 Species | n | 增温 Warming | 排水 Drainage | 增温 × 排水 Warming × Drainage | ||||
|---|---|---|---|---|---|---|---|---|
| 系数 Coefficient | 显著性 Significance | 系数 Coefficient | 显著性 Significance | 系数 Coefficient | 显著性 Significance | |||
| 圆锥薹草 Carex diandra | 高度 Height | 13 | 4.073 | 0.085 | 13.212 | 0.082 | -12.960 | 0.007 |
| 盖度 Coverage | 13 | -27.000 | 0.108 | -11.583 | 0.725 | 0.833 | 0.974 | |
| 频度 Frequency | 13 | -14.015 | 0.385 | -18.246 | 0.671 | -30.882 | 0.268 | |
| 地上生物量 Above-ground biomass | 13 | -2.692 | 0.362 | -1.731 | 0.774 | -0.322 | 0.934 | |
| 开花初始期 First flowering date | 7 | - | - | 8.000 | 0.210 | - | - | |
| 开花结束期 Last flowering date | 7 | - | - | 10.250 | 0.009 | - | - | |
| 开花峰值期 Flowering peak date | 7 | - | - | 11.167 | 0.076 | - | - | |
| 开花持续时间 Flowering duration | 7 | - | - | 2.250 | 0.727 | - | - | |
| 最大开花数量 Maximum flowering number | 7 | - | - | -13.167 | 0.496 | - | - | |
| 水葡萄茶藨子 Ribes procumbens | 高度 Height | 16 | 2.070 | 0.512 | -2.947 | 0.550 | 3.880 | 0.352 |
| 盖度 Coverage | 16 | -5.000 | 0.741 | -17.333 | 0.468 | 34.000 | 0.105 | |
| 频度 Frequency | 16 | -5.200 | 0.785 | -13.867 | 0.641 | 26.533 | 0.298 | |
| 地上生物量 Above-ground biomass | 16 | -6.507 | 0.263 | -8.738 | 0.299 | 10.792 | 0.138 | |
| 开花初始期 First flowering date | 15 | 1.727 | 0.449 | 1.382 | 0.799 | -2.524 | 0.604 | |
| 开花结束期 Last flowering date | 15 | 3.788 | 0.097 | 3.164 | 0.542 | 3.595 | 0.434 | |
| 开花峰值期 Flowering peak date | 15 | 5.576 | 0.104 | 4.327 | 0.581 | 4.571 | 0.512 | |
| 开花持续时间 Flowering duration | 15 | 2.061 | 0.418 | 1.782 | 0.768 | 6.119 | 0.247 | |
| 最大开花数量 Maximum flowering number | 15 | 1.424 | 0.813 | 3.073 | 0.832 | 14.548 | 0.252 | |
| 杜香 Ledum palustre | 高度 Height | 29 | 6.000 | 0.040 | 1.223 | 0.870 | -10.108 | 0.024 |
| 盖度 Coverage | 29 | 19.250 | 0.025 | -11.590 | 0.541 | -21.502 | 0.089 | |
| 频度 Frequency | 29 | 3.500 | 0.591 | -3.088 | 0.832 | -23.450 | 0.023 | |
| 地上生物量 Above-ground biomass | 29 | 12.999 | 0.249 | -20.981 | 0.579 | 5.287 | 0.824 | |
| 开花初始期 First flowering date | 16 | 4.088 | 0.177 | 3.514 | 0.642 | 6.933 | 0.241 | |
| 开花结束期 Last flowering date | 16 | -1.546 | 0.642 | -0.139 | 0.987 | -0.039 | 0.996 | |
| 开花峰值期 Flowering peak date | 16 | 6.097 | 0.043 | 3.042 | 0.671 | -3.051 | 0.457 | |
| 开花持续时间 Flowering duration | 16 | -5.634 | 0.243 | -3.653 | 0.763 | -7.315 | 0.470 | |
| 最大开花数量 Maximum flowering number | 16 | 0.838 | 0.982 | -24.736 | 0.796 | -52.318 | 0.493 | |
| 齿叶风毛菊 Saussurea neoserrata | 高度 Height | 11 | -9.412 | 0.614 | -12.858 | 0.759 | 5.733 | 0.828 |
| 盖度 Coverage | 11 | -1.914 | 0.671 | -1.052 | 0.918 | -1.315 | 0.837 | |
| 频度 Frequency | 11 | 4.000 | 0.757 | 4.667 | 0.866 | -18.667 | 0.336 | |
| 地上生物量 Above-ground biomass | 11 | -3.670 | 0.186 | -3.098 | 0.438 | 3.361 | 0.326 | |
| 开花初始期 First flowering date | 8 | 3.464 | 0.623 | -17.036 | 0.289 | -15.500 | 0.311 | |
| 开花结束期 Last flowering date | 8 | 3.143 | 0.447 | -9.857 | 0.287 | -1.000 | 0.916 | |
| 开花峰值期 Flowering peak date | 8 | 3.714 | 0.448 | -12.284 | 0.265 | -12.201 | 0.235 | |
| 开花持续时间 Flowering duration | 8 | -0.321 | 0.943 | 7.179 | 0.477 | 14.500 | 0.105 | |
| 最大开花数量 Maximum flowering number | 8 | -3.929 | 0.344 | -2.929 | 0.734 | 3.000 | 0.746 | |
附录I 广义线性混合模型统计结果
Supplement I Results of GLME model
| 物种 Species | n | 增温 Warming | 排水 Drainage | 增温 × 排水 Warming × Drainage | ||||
|---|---|---|---|---|---|---|---|---|
| 系数 Coefficient | 显著性 Significance | 系数 Coefficient | 显著性 Significance | 系数 Coefficient | 显著性 Significance | |||
| 圆锥薹草 Carex diandra | 高度 Height | 13 | 4.073 | 0.085 | 13.212 | 0.082 | -12.960 | 0.007 |
| 盖度 Coverage | 13 | -27.000 | 0.108 | -11.583 | 0.725 | 0.833 | 0.974 | |
| 频度 Frequency | 13 | -14.015 | 0.385 | -18.246 | 0.671 | -30.882 | 0.268 | |
| 地上生物量 Above-ground biomass | 13 | -2.692 | 0.362 | -1.731 | 0.774 | -0.322 | 0.934 | |
| 开花初始期 First flowering date | 7 | - | - | 8.000 | 0.210 | - | - | |
| 开花结束期 Last flowering date | 7 | - | - | 10.250 | 0.009 | - | - | |
| 开花峰值期 Flowering peak date | 7 | - | - | 11.167 | 0.076 | - | - | |
| 开花持续时间 Flowering duration | 7 | - | - | 2.250 | 0.727 | - | - | |
| 最大开花数量 Maximum flowering number | 7 | - | - | -13.167 | 0.496 | - | - | |
| 水葡萄茶藨子 Ribes procumbens | 高度 Height | 16 | 2.070 | 0.512 | -2.947 | 0.550 | 3.880 | 0.352 |
| 盖度 Coverage | 16 | -5.000 | 0.741 | -17.333 | 0.468 | 34.000 | 0.105 | |
| 频度 Frequency | 16 | -5.200 | 0.785 | -13.867 | 0.641 | 26.533 | 0.298 | |
| 地上生物量 Above-ground biomass | 16 | -6.507 | 0.263 | -8.738 | 0.299 | 10.792 | 0.138 | |
| 开花初始期 First flowering date | 15 | 1.727 | 0.449 | 1.382 | 0.799 | -2.524 | 0.604 | |
| 开花结束期 Last flowering date | 15 | 3.788 | 0.097 | 3.164 | 0.542 | 3.595 | 0.434 | |
| 开花峰值期 Flowering peak date | 15 | 5.576 | 0.104 | 4.327 | 0.581 | 4.571 | 0.512 | |
| 开花持续时间 Flowering duration | 15 | 2.061 | 0.418 | 1.782 | 0.768 | 6.119 | 0.247 | |
| 最大开花数量 Maximum flowering number | 15 | 1.424 | 0.813 | 3.073 | 0.832 | 14.548 | 0.252 | |
| 杜香 Ledum palustre | 高度 Height | 29 | 6.000 | 0.040 | 1.223 | 0.870 | -10.108 | 0.024 |
| 盖度 Coverage | 29 | 19.250 | 0.025 | -11.590 | 0.541 | -21.502 | 0.089 | |
| 频度 Frequency | 29 | 3.500 | 0.591 | -3.088 | 0.832 | -23.450 | 0.023 | |
| 地上生物量 Above-ground biomass | 29 | 12.999 | 0.249 | -20.981 | 0.579 | 5.287 | 0.824 | |
| 开花初始期 First flowering date | 16 | 4.088 | 0.177 | 3.514 | 0.642 | 6.933 | 0.241 | |
| 开花结束期 Last flowering date | 16 | -1.546 | 0.642 | -0.139 | 0.987 | -0.039 | 0.996 | |
| 开花峰值期 Flowering peak date | 16 | 6.097 | 0.043 | 3.042 | 0.671 | -3.051 | 0.457 | |
| 开花持续时间 Flowering duration | 16 | -5.634 | 0.243 | -3.653 | 0.763 | -7.315 | 0.470 | |
| 最大开花数量 Maximum flowering number | 16 | 0.838 | 0.982 | -24.736 | 0.796 | -52.318 | 0.493 | |
| 齿叶风毛菊 Saussurea neoserrata | 高度 Height | 11 | -9.412 | 0.614 | -12.858 | 0.759 | 5.733 | 0.828 |
| 盖度 Coverage | 11 | -1.914 | 0.671 | -1.052 | 0.918 | -1.315 | 0.837 | |
| 频度 Frequency | 11 | 4.000 | 0.757 | 4.667 | 0.866 | -18.667 | 0.336 | |
| 地上生物量 Above-ground biomass | 11 | -3.670 | 0.186 | -3.098 | 0.438 | 3.361 | 0.326 | |
| 开花初始期 First flowering date | 8 | 3.464 | 0.623 | -17.036 | 0.289 | -15.500 | 0.311 | |
| 开花结束期 Last flowering date | 8 | 3.143 | 0.447 | -9.857 | 0.287 | -1.000 | 0.916 | |
| 开花峰值期 Flowering peak date | 8 | 3.714 | 0.448 | -12.284 | 0.265 | -12.201 | 0.235 | |
| 开花持续时间 Flowering duration | 8 | -0.321 | 0.943 | 7.179 | 0.477 | 14.500 | 0.105 | |
| 最大开花数量 Maximum flowering number | 8 | -3.929 | 0.344 | -2.929 | 0.734 | 3.000 | 0.746 | |
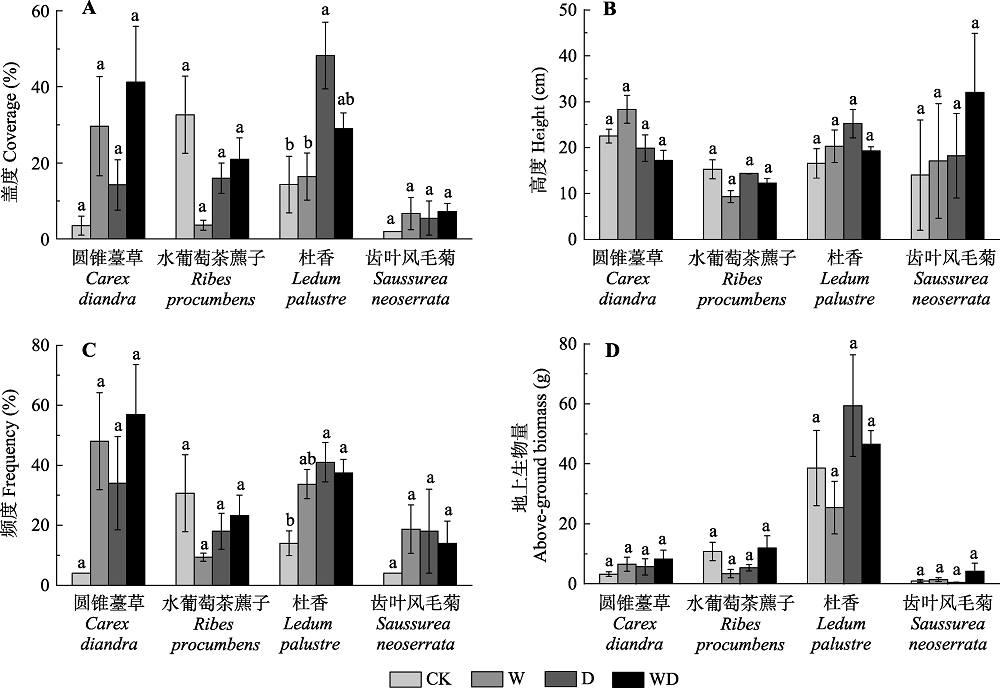
图3 模拟增温和排水处理对灌草层代表性物种的盖度(A)、高度(B)、频度(C)及地上生物量(D)的影响(平均值±标准误差)。相同物种内不同小写字母表示处理间差异显著(p < 0.05)。CK、W、D和WD分别代表对照、增温、排水和增温+排水处理。
Fig. 3 Simulated warming and drainage effects on coverage (A), height (B), frequency (C) and above-ground biomass (D) of the shrub-grass typical species (mean ± SE). Different small letters in the same species indicated significant difference among different treatments (p < 0.05). CK, control; W, warming; D, drainage; WD, warming + drainage.
| [1] |
Abbott BW, Jones JB, Schuur EAG, Iii FSC, Bowden WB, Bretharte MS, Epstein HE, Flannigan MD, Harms TK, Hollingsworth TN ( 2016). Biomass offsets little or none of permafrost carbon release from soils, streams, and wildfire: An expert assessment. Environmental Research Letters, 11, 034014. DOI: 10.1088/17489326/11/3/034014.
DOI URL |
| [2] |
Ackerly DD ( 2003). Community assembly, niche conservatism, and adaptive evolution in changing environments. International Journal of Plant Sciences, 164(Suppl. 3), S165-S184.
DOI URL |
| [3] |
Amano T, Smithers RJ, Sparks TH, Sutherland WJ ( 2010). A 250-year index of first flowering dates and its response to temperature changes. Proceedings of the Royal society B, 277, 2451-2457.
DOI URL PMID |
| [4] |
Anderson JT, Inouye DW, McKinney AM, Colautti RI, Mitchell-Olds T ( 2012). Phenotypic plasticity and adaptive evolution contribute to advancing flowering phenology in response to climate change. Proceedings of the Royal Society of London B: Biological Sciences, 279, 3843-3852.
DOI URL PMID |
| [5] |
Ausín I, Alonso-Blanco C, Martínez-Zapater JM ( 2005). Environmental regulation of flowering. International Journal of Developmental Biology, 49, 689-705.
DOI URL |
| [6] |
Badeck FW, Bondeau A, Böttcher K, Doktor D, Lucht W, Schaber J, Sitch S ( 2004). Responses of spring phenology to climate change. New Phytologist, 162, 295-309.
DOI URL |
| [7] | Brouwer R ( 1983). Functional equilibrium: Sense or nonsense. Netherlands Journal of Agricultural Science, 31, 335-348. |
| [8] |
Brown DRN, Torre JM, Douglas TA, Romanovsky VE, Knut K, Christopher H, Euskirchen ES, Ruess RW ( 2015). Interactive effects of wildfire and climate on permafrost degradation in Alaskan lowland forests. Journal of Geophysical Research Biogeosciences, 120, 1619-1637.
DOI URL |
| [9] |
CaraDonna PJ, Iler AM, Inouye DW ( 2014). Shifts in flowering phenology reshape a subalpine plant community. Proceedings of the National Academy of Sciences of the United States of America, 111, 4916-4921.
DOI URL |
| [10] |
Cleland EE, Allen JM, Crimmins TM, Dunne JA, Pau S, Travers SE, Zavaleta ES, Wolkovich EM ( 2012). Phenological tracking enables positive species responses to climate change. Ecology, 93, 1765-1771.
DOI URL PMID |
| [11] |
Cleland EE, Chiariello NR, Loarie SR, Mooney HA, Field CB ( 2006). Diverse responses of phenology to global changes in a grassland ecosystem. Proceedings of the National Academy of Sciences of the United States of America, 103, 13740-13744.
DOI URL PMID |
| [12] |
Cleland EE, Chuine I, Menzel A, Mooney HA, Schwartz MD ( 2007). Shifting plant phenology in response to global change. Trends in Ecology and Evolution, 22, 357-365.
DOI URL PMID |
| [13] |
Dorji T, Totland O, Moe SR, Hopping KA, Pan J, Klein JA ( 2013). Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet. Global Change Biology, 19, 459-472.
DOI URL PMID |
| [14] |
Dudgeon SR, Steneck RS, Davison IR, Vadas RL ( 1999). Coexistence of similar species in a space-limited intertidal zone. Ecological Monographs, 69, 331-352.
DOI URL |
| [15] |
Forrest J, Miller-Rushing AJ ( 2010). Toward a synthetic understanding of the role of phenology in ecology and evolution. Philosophical Transactions of the Royal Society B, 365, 3101-3112.
DOI URL PMID |
| [16] | Galen C, Sheery RA, Carroll AB ( 1999). Are flowers physiolosical sinks or faucets? Costs ans correlates of water use by flowers of Polemonium viscosum. Oecologia, 118, 461-470. |
| [17] |
Gugger S, Kesselring H, Stöcklin J, Hamann E ( 2015). Lower plasticity exhibited by high-versus mid-elevation species in their phenological responses to manipulated temperature and drought. Annals of Botany, 116, 953-962.
DOI URL PMID |
| [18] |
Hulme PE ( 2011). Contrasting impacts of climate-driven flowering phenology on changes in alien and native plant species distributions. New Phytologist, 189, 272-281.
DOI URL PMID |
| [19] |
IPCC ( Intergovernmental Panel on Climate Change) ( 2013). The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, New York.
DOI URL |
| [20] |
Jorgenson MT, Racine CH, Walters JC, Osterkamp TE ( 2001). Permafrost degradation and ecological changes associated with a warming climate in central Alaska. Climatic Change, 48, 551-579.
DOI URL |
| [21] |
Kim Y, Tanaka N ( 2003). Effect of forest fire on the fluxes of CO2, CH4 and N2O in boreal forest soils, interior Alaska. Journal of Geophysical Research, 108, 8154. DOI: 10.1029/2001JD000663.
DOI URL |
| [22] |
Kochmer JP, Handel SN ( 1986). Constraints and competition in the evolution of flowering phenology. Ecological Monographs, 56, 303-325.
DOI URL |
| [23] |
Li X, Jiang L, Meng F, Wang S, Niu H, Iler AM, Duan J, Zhang Z, Luo C, Cui S, Zhang L, Li Y, Wang Q, Zhou Y, Bao X, Doriji T, Li Y, Peñuelas J, Du M, Zhao X, Zhao L, Wang G ( 2016). Responses of sequential and hierarchical phenological events to warming and cooling in alpine meadows. Nature Communications, 7, 12489. DOI: 10.1038/ncomms12489.
DOI URL PMID |
| [24] |
Meng F, Cui S, Wang S, Duan J, Jiang L, Zhang Z, Luo C, Wang Q, Zhou Y, Li X ( 2016). Changes in phenological sequences of alpine communities across a natural elevation gradient. Agricultural and Forest Meteorology, 224, 11-16.
DOI URL |
| [25] |
Meng FD, Jiang LL, Zhang ZH, Cui SJ, Duan JC, Wang SP, Luo CY, Wang Q, Zhou Y, Li XE, Zhang LR, Li BW, Dorij T, Li YN, Du MY ( 2017). Changes in flowering functional group affect responses of community phenological sequences to temperature change. Ecology, 98, 734-740.
DOI URL PMID |
| [26] |
Menzel A, Seifert H, Eatrella N ( 2011). Effects of recent warm and cold spells on European plant phenology. International Journal of Biometeorology, 55, 921-932.
DOI URL PMID |
| [27] |
Peñuelas J, Filella I ( 2001). Phenology: Reapones to a warming world. Science, 294, 793-795.
DOI URL PMID |
| [28] |
Peñuelas J, Filella I, Comas P ( 2002). Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Global Change Biology, 8, 531-544.
DOI URL |
| [29] |
Peñuelas J, Prieto P, Beier C, Cesaraccio C, de Angelis P, de Dato G, Emmett BA, Estiarte M, Garadnai J, Gorissen A ( 2007). Response of plant species richness and primary productivity in shrublands along a north-south gradient in Europe to seven years of experimental warming and drought: Reductions in primary productivity in the heat and drought year of 2003. Global Change Biology, 13, 2563-2581.
DOI URL |
| [30] |
Peñuelas J, Rutishauser T, Filella I ( 2009). Phenology feedbacks on climate change. Science, 324, 887-888.
DOI URL PMID |
| [31] |
Post ES, Pedersen C, Wilmers CC, Forchhammer MC ( 2008). Phenological sequences reveal aggregate life history response to climatic warming. Ecology, 89, 363-370.
DOI URL PMID |
| [32] |
Schemske DW, Willson MF, Melampy MN, Miller LJ, Verner L, Schemske KM, Best LB ( 1978). Flowering ecology of some spring woodland herbs. Ecology, 59, 351-366.
DOI URL |
| [33] |
Sherry RA, Zhou X, Gu S, Arnone JA, Schimel DS, Verburg PS, Wallace LL, Luo Y ( 2007). Divergence of reproductive phenology under climate warming. Proceedings of the National Academy of Sciences of the United States of America, 104, 198-202.
DOI URL |
| [34] |
Steinaker DF, Wilson SD, Peltzer DA ( 2010). Asynchronicity in root and shoot phenology in grass and woody plants. Global Change Biology, 16, 2241-2251.
DOI URL |
| [35] |
Sullivan PF, Welker JM ( 2005). Warming chambers stimulate early season growth of an arctic sedge: Results of a minirhizotron field study. Oecologia, 142, 616-626.
DOI URL PMID |
| [36] | Sun MX ( 2011). The Impacts on Soil Properities and Revegetation from Forest in Tahe Forest Region. PhD dissertation, Bejing Forestry University,Beijing. |
| [ 孙明学 ( 2011). 塔河林区林火对土壤性质与植被恢复的影响. 博士学位论文, 北京林业大学, 北京.] | |
| [37] |
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJ, Fromentin JM, Hoegh-Guldberg O, Bairlein F ( 2002). Ecological responses to recent climate change. Nature, 416, 389-395.
DOI URL PMID |
| [38] |
Wang S, Meng F, Duan J, Wang Y, Cui X, Piao S, Niu H, Xu G, Luo C, Zhang Z ( 2014 a). Asymmetric sensitivity of first flowering date to warming and cooling in alpine plants. Ecology, 95, 3387-3398.
DOI URL |
| [39] |
Wang S, Wang C, Duan J, Zhu X, Xu G, Luo C, Zhang Z, Meng F, Li Y, Du M ( 2014 b). Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau. Agricultural and Forest Meteorology, 189, 220-228.
DOI URL |
| [40] | Wang XG, Li XZ, He HS, Leng WF, Wen QC ( 2004). Postfire succession of larch forest in the northern slope of Daxinganling. Chinese Journal of Ecology, 23, 35-41. |
| [ 王绪高, 李秀珍, 贺红士, 冷文芳, 问青春 ( 2004). 大兴安岭北坡落叶松林火后植被演替过程研究. 生态学杂志, 23, 35-41.] | |
| [41] |
Wolkovich EM, Cook BI, Allen JM, Crimmins TM, Betancourt JL, Travers SE, Pau S, Regetz J, Davies TJ, Kraft NJ ( 2012). Warming experiments underpredict plant phenological responses to climate change. Nature, 485, 494-497.
DOI URL PMID |
| [42] | Wu RF, Huo ZG, Cao YF, Yan WX ( 2009). Phenophase change of typical herbaceous plants in Inner Mongolia in spring and its response to climate warming. Chinese Journal of Ecology, 28, 1470-1475. |
| [ 吴瑞芬, 霍治国, 曹艳芳, 闫伟兄 ( 2009). 内蒙古典型草本植物春季物候变化及其对气候变暖的响应. 生态学杂志, 28, 1470-1475.] | |
| [43] |
Ye X, Zhou HK, Liu GH, Yao BQ, Zhao XQ ( 2014). Responses of phenological characteristics of major plants to nutrient and water additions in Kobresia humilis alpine meadow. Chinese Journal of Plant Ecology, 38, 147-158.
DOI URL |
|
[ 叶鑫, 周华坤, 刘国华, 姚步青, 赵新全 ( 2014). 高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应. 植物生态学报, 38, 147-158.]
DOI URL |
|
| [44] |
Yu C, He HS, Hu Y, Bu R, Li X ( 2008). Historic and current fire regimes in the Great Xing’an Mountains, northeastern China: Implications for long-term forest management. Forest Ecology and Management, 254, 445-453.
DOI URL |
| [1] | 郭敏, 罗林, 梁进, 王彦杰, 赵春章. 冻融变化对西南亚高山森林优势种云杉和华西箭竹根区土壤理化性质与酶活性的影响[J]. 植物生态学报, 2023, 47(6): 882-894. |
| [2] | 魏瑶, 马志远, 周佳颖, 张振华. 模拟增温改变青藏高原植物繁殖物候及植株高度[J]. 植物生态学报, 2022, 46(9): 995-1004. |
| [3] | 魏春雪, 杨璐, 汪金松, 杨家明, 史嘉炜, 田大栓, 周青平, 牛书丽. 实验增温对陆地生态系统根系生物量的影响[J]. 植物生态学报, 2021, 45(11): 1203-1212. |
| [4] | 朱彪, 陈迎. 陆地生态系统野外增温控制实验的技术与方法[J]. 植物生态学报, 2020, 44(4): 330-339. |
| [5] | 李旭, 吴婷, 程严, 谭钠丹, 蒋芬, 刘世忠, 褚国伟, 孟泽, 刘菊秀. 南亚热带常绿阔叶林4个树种对增温的生理生态适应能力比较[J]. 植物生态学报, 2020, 44(12): 1203-1214. |
| [6] | 闫鹏飞, 展鹏飞, 肖德荣, 王燚, 余瑞, 刘振亚, 王行. 模拟增温及分解界面对茭草凋落物分解速率及叶际微生物结构和功能的影响[J]. 植物生态学报, 2019, 43(2): 107-118. |
| [7] | 张莉, 王根绪, 冉飞, 彭阿辉, 肖瑶, 杨阳, 杨燕. 模拟增温改变川西高山草甸优势植物繁殖物候序列特征[J]. 植物生态学报, 2018, 42(1): 20-27. |
| [8] | 管超, 张鹏, 李新荣. 腾格里沙漠东南缘生物结皮土壤呼吸对水热因子变化的响应[J]. 植物生态学报, 2017, 41(3): 301-310. |
| [9] | 朱军涛. 实验增温对藏北高寒草甸植物繁殖物候的影响[J]. 植物生态学报, 2016, 40(10): 1028-1036. |
| [10] | 徐冰鑫, 胡宜刚, 张志山, 陈永乐, 张鹏, 李刚. 模拟增温对荒漠生物土壤结皮-土壤系统CO2、CH4和N2O通量的影响[J]. 植物生态学报, 2014, 38(8): 809-820. |
| [11] | 熊沛, 徐振锋, 林波, 刘庆. 岷江上游华山松林冬季土壤呼吸对模拟增温的短期响应[J]. 植物生态学报, 2010, 34(12): 1369-1376. |
| [12] | 徐振锋, 尹华军, 赵春章, 曹刚, 万名利, 刘庆. 陆地生态系统凋落物分解对全球气候变暖的响应[J]. 植物生态学报, 2009, 33(6): 1208-1219. |
| [13] | 李志勇, 陈建军, 王彦辉, 于澎涛, 杜士才, 何萍, 段健. 重庆酸雨区人工木荷林对土壤化学性质的影响[J]. 植物生态学报, 2008, 32(3): 632-638. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19