
植物生态学报 ›› 2005, Vol. 29 ›› Issue (6): 976-984.DOI: 10.17521/cjpe.2005.0130
孟令曾1,2( ), 张教林1,2, 曹坤芳1, 许再富1
), 张教林1,2, 曹坤芳1, 许再富1
收稿日期:2004-12-26
接受日期:2005-04-19
出版日期:2005-12-26
发布日期:2005-09-30
作者简介:E-mail:mlz@xtbg.org.en
基金资助:
MENG Ling-Zeng1,2(), ZHANG Jiao-Lin1,2, CAO Kun-Fang1, XU Zai-Fu1
Received:2004-12-26
Accepted:2005-04-19
Online:2005-12-26
Published:2005-09-30
摘要:
龙脑香科植物是东南亚热带雨林冠层的优势树种,在生态和经济上具有重要地位,而我国西南地区被认为是龙脑香科植物分布的最北端。该文于2004年雨季测定了西双版纳热带植物园迁地保护区内引种的4种国产龙脑香科植物:望天树(Parashorea chinensis)、云南龙脑香(Dipterocarpus retusus)、版纳青梅(Vatica xishuangbannaensis)和海南坡垒(Hopea hainanensis)冠层叶片的光响应曲线、光合速率的日进程、叶绿素荧光参数、叶绿素含量及叶片平均面积、气孔保卫细胞长度和分布状况等。结果表明,4种植物的最大光合速率(Pmax)(7.5~18.1 μmol·m -2·s-1,用单位叶面积表示;89.1~150.8 nmol·g-1 DW·s-1,用单位干重表示)、暗呼吸速率(Rd)、光补偿点(LSP)、光饱和点(LCP)、表观量子效率和叶片特征差异显著。在正午时4个树种均出现光合作用显著降低的现象,其中海南坡垒光合速率下降最少。虽然高的叶片温度对光合作用有一定的影响,但正午光合作用的下降主要是由于正午高的叶片-空气的水蒸气压力差(LAVPD)引起的气孔关闭造成的。4个树种正午的光系统Ⅱ线性电子传递的量子效率ΦPSⅡ显著降低,表明遭受了强烈的光抑制。云南龙脑香、版纳青梅和海南坡垒的非光化学猝灭系数(NPQ)在正午时升高的幅度最大,而望天树的NPQ上升较少,表明热耗散是前3个树种的主要光保护机制。中午前后望天树的表观电子传递速率(ETR)一直保持在很高的水平,但其光合速率显著降低,表明大量的电子被分配到光呼吸上,也说明望天树主要通过光呼吸来保护光合机构。
孟令曾, 张教林, 曹坤芳, 许再富. 迁地保护的4种龙脑香冠层叶光合速率和叶绿素荧光参数的日变化. 植物生态学报, 2005, 29(6): 976-984. DOI: 10.17521/cjpe.2005.0130
MENG Ling-Zeng, ZHANG Jiao-Lin, CAO Kun-Fang, XU Zai-Fu. DIURNAL CHANGES OF PHOTOSYNTHETIC CHARACTERISTICS AND CHLOROPHYLL FLUORESCENCE IN CANOPY LEAVES OF FOUR DIPTOCARP SPECIES UNDER EX-SITU CONSERVATION. Chinese Journal of Plant Ecology, 2005, 29(6): 976-984. DOI: 10.17521/cjpe.2005.0130
| 树种 Species | 云南龙脑香 Dipterocarpus retusus | 海南坡垒 Hopea hainanensis | 望天树 Parashorea chinensis | 版纳青梅 Vatica xishuangbannaensis | |||
|---|---|---|---|---|---|---|---|
| 样木平均高度 Height of sampled trees (m) | 21 (45)* | 18 (25) | 17 (60) | 15 (40) | |||
| 样木平均胸径 DBH of sampled trees (cm) | 25.3 | 22.3 | 19.0 | 14.0 | |||
| 中国分布地区和海拔 Regions and altitude distributed in China | 云南盈江, 西藏墨脱(600~1 000 m) Yingjiang in Yunnan, Motuo in Tibet (600-1 000 m) | 海南崖县、琼中(400~800 m) Yaxian, Qiongzhong in Hainan (400-800 m) | 云南补蚌, 广西那坡 (700~950 m) Bubeng in Yunnan, Napo in Guangxi (700-950 m) | 云南补蚌, 广西那坡 (800~1 100 m) Bubeng in Yunnan, Napo in Guangxi (800-1 100 m) | |||
表1 4个树种的基本信息
Table 1 The general information of the four species selected
| 树种 Species | 云南龙脑香 Dipterocarpus retusus | 海南坡垒 Hopea hainanensis | 望天树 Parashorea chinensis | 版纳青梅 Vatica xishuangbannaensis | |||
|---|---|---|---|---|---|---|---|
| 样木平均高度 Height of sampled trees (m) | 21 (45)* | 18 (25) | 17 (60) | 15 (40) | |||
| 样木平均胸径 DBH of sampled trees (cm) | 25.3 | 22.3 | 19.0 | 14.0 | |||
| 中国分布地区和海拔 Regions and altitude distributed in China | 云南盈江, 西藏墨脱(600~1 000 m) Yingjiang in Yunnan, Motuo in Tibet (600-1 000 m) | 海南崖县、琼中(400~800 m) Yaxian, Qiongzhong in Hainan (400-800 m) | 云南补蚌, 广西那坡 (700~950 m) Bubeng in Yunnan, Napo in Guangxi (700-950 m) | 云南补蚌, 广西那坡 (800~1 100 m) Bubeng in Yunnan, Napo in Guangxi (800-1 100 m) | |||
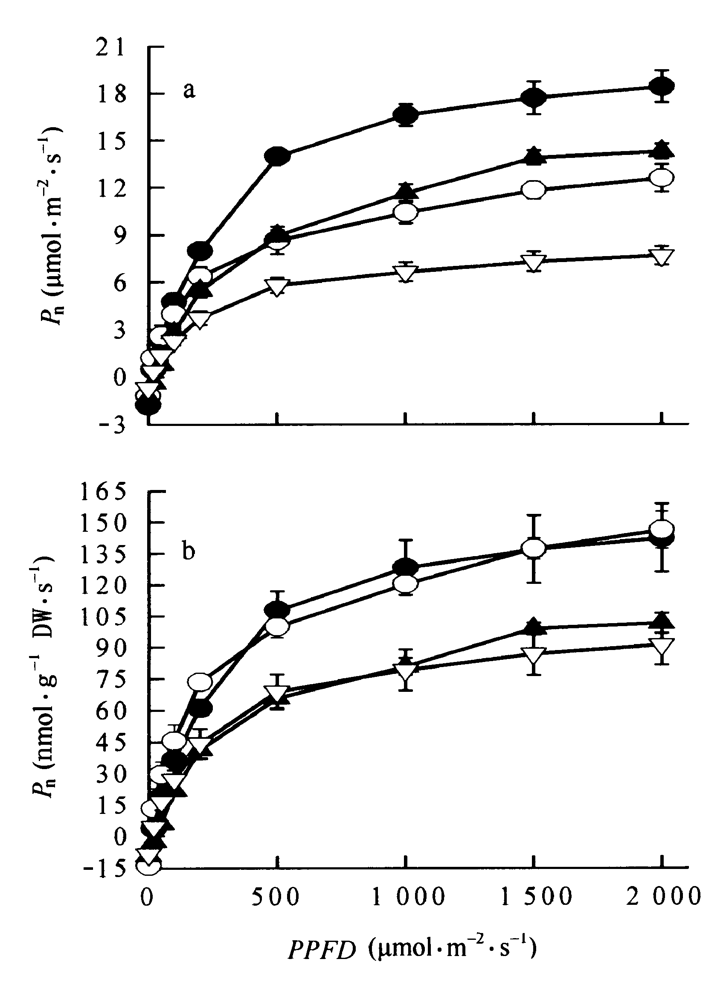
图1 4个树种冠层叶光合作用的光响应曲线 a: 以单位叶面积表示 On area basis b: 以单位干重表示 On mass basis ●云南龙脑香Dipterocarpus retusus ○望天树Parashorea chinensis ▲海南坡垒Hopea hainanensis ▽版纳青梅Vatica xishuangbannaensis 数据点为平均值±标准误 Data were means±SE (n=3-4) Pn: 净光合速率 Net photosynthesis rate PPFD: 光通量密度 Photon flux density
Fig.1 The light-photosynthetic response curves of canopy leaves in four species
| 树种 Species | 云南龙脑香 Dipterocarpus retusus | 海南坡垒 Hopea hainanensis | 望天树 Parashorea chinensis | 版纳青梅 Vatica xishuangbannaensis |
|---|---|---|---|---|
| 最大净光合速率 Maximal net photosynthetic rate (Pmax) | ||||
| 以单位叶面积表示 On area basis (μmol·m-2·s-1) | 18.10±1.13a | 14.43±0.52b | 12.97±0.93b | 7.50±0.59c |
| 以单位干重表示 On dry weight (nmol ·g -1 DW·s-1) | 140.10±16.93a | 101.79±2.52b | 150.82±11.32a | 89.08±8.27b |
| 暗呼吸速率 Dark respiration rate (Rd) | ||||
| 以单位叶面积表示 On area basis (μmol·m-2·s-1) | 2.08±0.15a | 1.35±0.09b | 1.22±0.03b | 0.70±0.10c |
| 以单位干重表示 On dry weight (nmol ·g -1 DW·s-1) | 13.86±2.06a | 9.13±1.03a | 13.98±0.88a | 8.44±1.15a |
| 光饱和点 Light satuaration point (μmol·m-2·s-1) | 791±93b | 1184±109a | 1311±204a | 851±81b |
| 光补偿点 Light compensation point (μmol·m-2·s-1) | 15.8±0.9b | 30.4±2.6a | 10.3±2.0b | 11.6±1.8b |
| 表观量子效率 Apparent quantum yield (μmol·μmol-1) | 0.062±0.005a | 0.034±0.002b | 0.051±0.008ab | 0.031±0.003b |
表2 4个树种冠层叶片的气体交换参数
Table 2 Gas exchange parameters of the canopy leaves in four species
| 树种 Species | 云南龙脑香 Dipterocarpus retusus | 海南坡垒 Hopea hainanensis | 望天树 Parashorea chinensis | 版纳青梅 Vatica xishuangbannaensis |
|---|---|---|---|---|
| 最大净光合速率 Maximal net photosynthetic rate (Pmax) | ||||
| 以单位叶面积表示 On area basis (μmol·m-2·s-1) | 18.10±1.13a | 14.43±0.52b | 12.97±0.93b | 7.50±0.59c |
| 以单位干重表示 On dry weight (nmol ·g -1 DW·s-1) | 140.10±16.93a | 101.79±2.52b | 150.82±11.32a | 89.08±8.27b |
| 暗呼吸速率 Dark respiration rate (Rd) | ||||
| 以单位叶面积表示 On area basis (μmol·m-2·s-1) | 2.08±0.15a | 1.35±0.09b | 1.22±0.03b | 0.70±0.10c |
| 以单位干重表示 On dry weight (nmol ·g -1 DW·s-1) | 13.86±2.06a | 9.13±1.03a | 13.98±0.88a | 8.44±1.15a |
| 光饱和点 Light satuaration point (μmol·m-2·s-1) | 791±93b | 1184±109a | 1311±204a | 851±81b |
| 光补偿点 Light compensation point (μmol·m-2·s-1) | 15.8±0.9b | 30.4±2.6a | 10.3±2.0b | 11.6±1.8b |
| 表观量子效率 Apparent quantum yield (μmol·μmol-1) | 0.062±0.005a | 0.034±0.002b | 0.051±0.008ab | 0.031±0.003b |
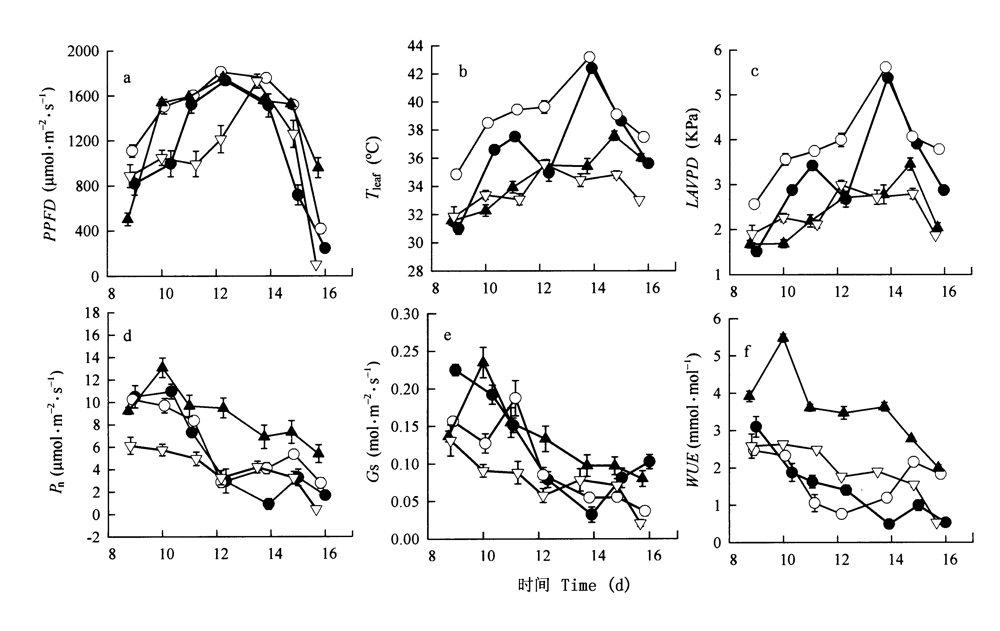
图2 4个树种净光合速率、气孔导度、水分利用效率及环境参数的日变化 ●云南龙脑香Dipterocarpus retusus ○望天树Parashorea chinensis ▲海南坡垒Hopea hainanensis ▽版纳青梅Vatica xishuangbannaensis 数据点为平均值±标准误 Data were means±SE (n=3-4) Pn、PPFD: 同图1 See Fig.1 Gs: 气孔导度 Stomatal conductance WUE: 水分利用效率 Water use efficiency LAVPD: 叶片到空气的水气压差 Leaf-to-air vapor pressure deficit
Fig.2 The diurnal changes in net photosynthetic rate (Pn), stomatal conductance (Gs), water use efficiency (WUE) and environmental parameters of four dipterocarp species
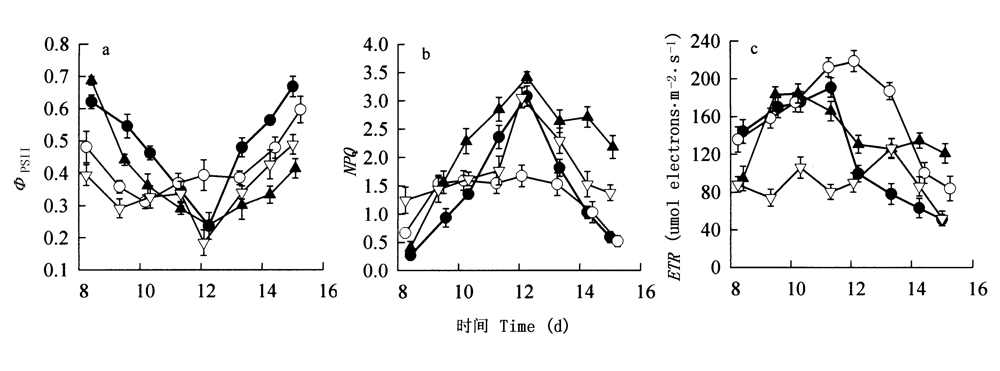
图3 4个树种叶绿素荧光参数的日变化 ●云南龙脑香Dipterocarpus retusus ○望天树Parashorea chinensis ▲海南坡垒Hopea hainanensis ▽版纳青梅Vatica xishuangbannaensis 数据点为平均值±标准误 Data were means±SE (n=3-4) ΦPSⅡ: 光系统II下的光量子效率 Quantum yields of PSII NPQ: 非光化学猝灭 Nonphotochemical quenching rate ETR: 电子传递速率 Electron transport rate
Fig.3 The diurnal changes in parameters of chlorophyll fluorescence in four dipterocarp species
| 树种 Species | 云南龙脑香 Dipterocarpus retusus | 海南坡垒 Hopea hainanensis | 望天树 Parashorea chinensis | 版纳青梅 Vatica xishuangbannaensis |
|---|---|---|---|---|
| 正午水势 Midday leaf water potential (MPa) | -0.058±0.003b | -0.071±0.007a | -0.094±0.012a | -0.072±0.004ab |
| 相对含水量 Relative water content (%) | 98.0±0.4a | 99.6±0.3a | 93.9±0.6b | 98.8±0.2a |
| 叶绿素含量 Total chl content (mg·g-1 FW) | 1.39±0.29b | 1.97±0.17a | 2.46±0.37a | 1.53±0.12b |
| 叶绿素a/b比值 Ratios of chl a/b | 2.71±0.05a | 2.50±0.07ab | 2.20±0.17b | 2.61±0.08a |
| 比叶面积 Specific leaf area (cm2·g-1) | 76.85±5.02b | 72.23±2.70b | 116.65±7.20a | 118.40±2.90a |
| 气孔密度 Stomatal density (mm-2) | 89.5±1.79c | 90.0±0.19c | 105.2±1.89b | 158.6±1.79a |
| 保卫细胞长度 Guard cell length (μm) | 25.41±0.69a | 22.10±0.47b | 21.96±0.23b | 19.77±0.50c |
| 平均叶面积 Average leaf area (cm2) | 403.8±26.7a | 66.6±2.5b | 47.6±2.1c | 42.9±2.1c |
表3 4个树种冠层叶片的叶片水势、叶绿素含量和叶片特征
Table 3 Leaf water potential, chlorophyll content and leaf traits in four dipterocarp species
| 树种 Species | 云南龙脑香 Dipterocarpus retusus | 海南坡垒 Hopea hainanensis | 望天树 Parashorea chinensis | 版纳青梅 Vatica xishuangbannaensis |
|---|---|---|---|---|
| 正午水势 Midday leaf water potential (MPa) | -0.058±0.003b | -0.071±0.007a | -0.094±0.012a | -0.072±0.004ab |
| 相对含水量 Relative water content (%) | 98.0±0.4a | 99.6±0.3a | 93.9±0.6b | 98.8±0.2a |
| 叶绿素含量 Total chl content (mg·g-1 FW) | 1.39±0.29b | 1.97±0.17a | 2.46±0.37a | 1.53±0.12b |
| 叶绿素a/b比值 Ratios of chl a/b | 2.71±0.05a | 2.50±0.07ab | 2.20±0.17b | 2.61±0.08a |
| 比叶面积 Specific leaf area (cm2·g-1) | 76.85±5.02b | 72.23±2.70b | 116.65±7.20a | 118.40±2.90a |
| 气孔密度 Stomatal density (mm-2) | 89.5±1.79c | 90.0±0.19c | 105.2±1.89b | 158.6±1.79a |
| 保卫细胞长度 Guard cell length (μm) | 25.41±0.69a | 22.10±0.47b | 21.96±0.23b | 19.77±0.50c |
| 平均叶面积 Average leaf area (cm2) | 403.8±26.7a | 66.6±2.5b | 47.6±2.1c | 42.9±2.1c |
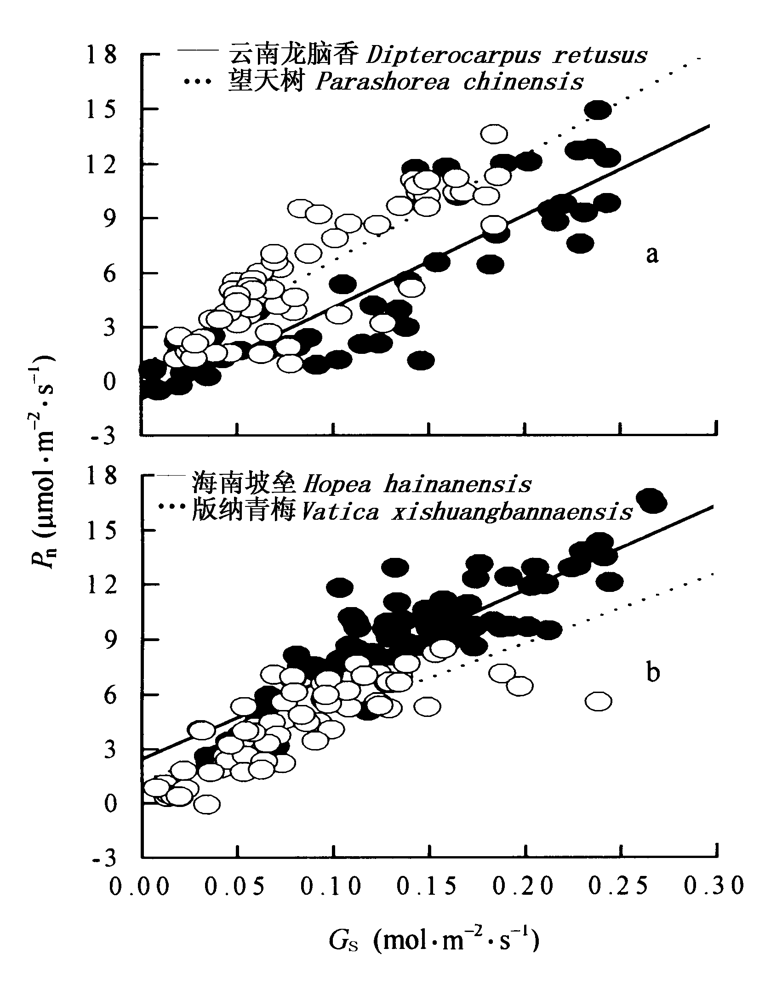
图4 4个树种净光合速率和气孔导度的相关性 数据为一天的所有测定值 Data were pooled from all measurements during a day 在图a和b中黑点分别代表云南龙脑香和海南坡垒, 白点分别代表望天树和版纳青梅 Black dot represents Dipterocarpus retusus and Hopea hainanensis, white dot represents Parashorea chinensis and Vatica xishuangbannaensis 4个种的回归方程分别是 The regression equations of four species are Pn=50.74Gs-0.96 R2=0.77 p<0.001n=48 Dipterocarpus retusus Pn=58.14Gs+0.84 R2=0.72 p<0.001n=60 Parashorea chinensis Pn=46.33Gs+2.45 R2=0.79 p<0.001n=86 Hopea hainanensis Pn=37.88Gs+1.21 R2=0.63 p<0.001n=71 Vatica xishuangbannaensis Pn、Gs: 同图2 See Fig.2
Fig.4 The relationship between Pn and Gs in four dipterocarp species
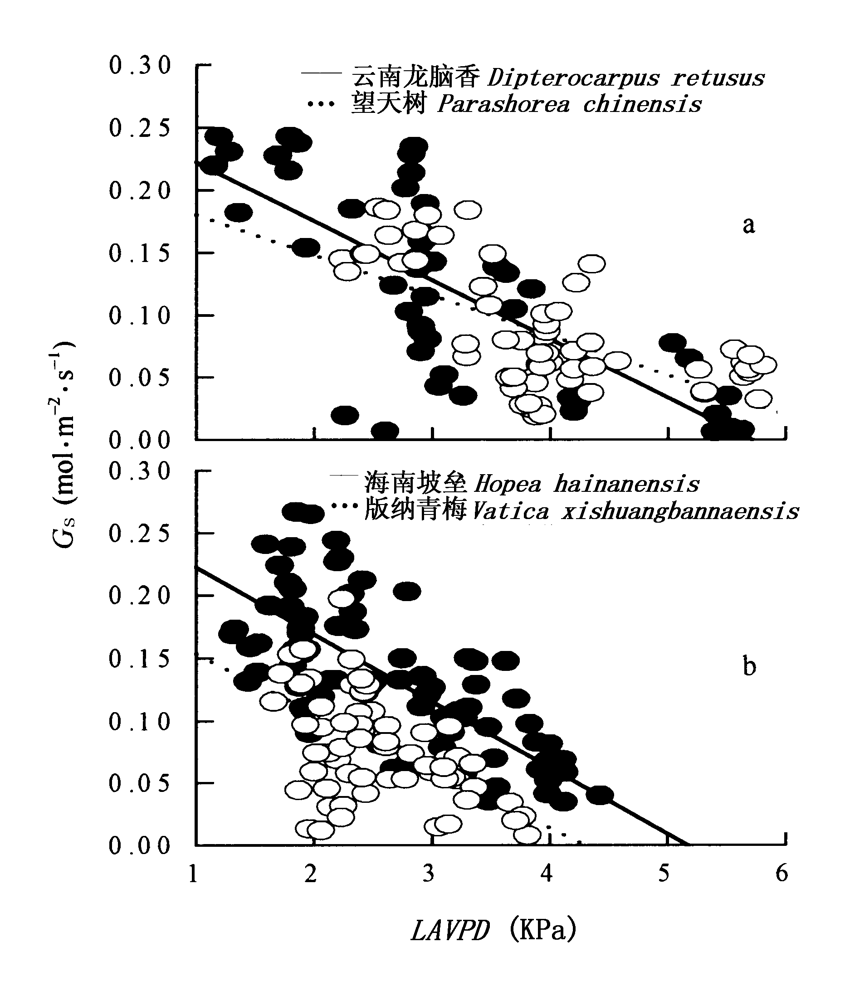
图5 4个树种气孔导度和叶片水蒸气压差的相关性 数据为一天的所有测定值 Data were pooled from all measurements during a day 在图a和b中黑点分别代表云南龙脑香和海南坡垒, 白点分别代表望天树和版纳青梅 Black dot represents Dipterocarpus retusus and Hopea hainanensis, white dot represents Parashorea chinensis and Vatica xishuangbannaensis 4个种的回归方程分别是 The regression equations of four species are Gs=-0.05LAVPD+0.27 R2=0.55 p<0.001n=48 Dipterocarpus retusus Gs=-0.03LAVPD+0.21 R2=0.39 p<0.001n=60 Parashorea chinensis Gs=-0.05LAVPD+0.28 R2=0.51 p<0.001n=86 Hopea hainanensis Gs=-0.05LAVPD+0.20 R2=0.37 p<0.001n=71Vatica xishuangbannaensis Gs、LAVPD: 同图2 See Fig.2
Fig.5 The relationship between LAVPD and Gs in four dipterocarp species
| [1] | Airy Shaw HK (1973). A Dictionary of the Flowering Plants and Ferns. 1st edn. Cambridge University Press, Cambridge,375-376. |
| [2] |
Al-Khatib K, Paulsen GM (1989). Enhancement of thermal injury to photosynthesis in wheat plants and thylakoids by high light intensity. Plant Physiology, 90,1041-1048.
URL PMID |
| [3] | Aylett GP (1985). Irradiance interception, leaf conductance and photosynthesis in Jamaican upper mountain rain forest trees. Photosynthetica, 19,323-337. |
| [4] | Bassow SL, Bazzaz FA (1998). How environmental conditions affect canopy leaf-level photosynthesis in four deciduous tree species. Ecology, 79,2660-2675. |
| [5] |
Bilger W, Bjørkman O (1990). Role of the xanthophylls cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in Hedera canariensis. Photosynthesis Research, 25,173-185.
DOI URL PMID |
| [6] | Brodribb TJ, Holbrook NM (2004). Diurnal depression of leaf hydraulic conductance in a tropical tree species. Plant, Cell and Environment, 27,820-827. |
| [7] | Cao KF, Booth EW (2001). Leaf anatomical structure and photosynthetic induction for seedlings of five dipterocarp species under contrasting light conditions in a Bornean heath forest. Journal of Tropical Ecology, 17,163-175. |
| [8] | Cao KF (2000). Water relations and gas exchange of tropical saplings during a prolonged drought in a Bornean heath forest, with reference to root architecture. Journal of Tropical Ecology, 16,101-116. |
| [9] | Demmig-Adams B, Adams WW Ⅲ (1992). Photoprotection and other responses of plants to high light stress. Annual Review of Plant Physiology and Plant Molecular Biology, 43,599-626. |
| [10] | Epron D (1997). Effects of drought on photosynthesis and on the thermotolerance of photosystem II in seedlings of cedar (Cedrus atlantica and Clibani). Journal of Experimental Botany, 48,1835-1841. |
| [11] | Foyer GH, Lelandais M, Kunert KJ (1994). Photooxidative stress in plants. Physiologia Plantarum, 92,696-717. |
| [12] |
Franco AC, Lüttge U (2002). Midday depression in savanna trees: coordinated adjustments in photo-chemical efficiency, photorespiration, CO 2 assimilation and water use efficiency. Oecologia, 131,356-365.
DOI URL PMID |
| [13] | Genty B, Briantais JM, Baker NR (1989). The relationship between the quantum yield of photosunthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta, 990,87-92. |
| [14] | Grace J, Okali DUU, Fasehun FE (1982). Stomatal conductance of two tropical trees during the wet season in Nigeria. Journal of Applied Ecology, 19,659-670. |
| [15] | Iio AH, Fukasawa YN, Kakubari Y (2004). Stomatal closure induced by high vapor pressure deficit limited midday photosynthesis at the canopy top of Fagus crenata Blume on Naeba mountain in Japan. Trees, 18,510-117. |
| [16] |
Ishida A, Toma T, Marjenah (1999). Limitation of leaf carbon gain by stomatal and photochemical processes in the top canopy of Macaranga conifera, a tropical pioneer tree. Tree Physiology, 19,467-473.
URL PMID |
| [17] |
Ishida A, Toma T, Matsumoto Y, Yap SK, Maruyama Y (1996). Diurnal changes in leaf gas exchange characteristics in the uppermost canopy of a rain forest tree, ( Dryobalanops aromatica) Gaertn. f. Tree Physiology, 16,779-785.
URL PMID |
| [18] | Johnston M, Grof CPL, Brownell PF (1984). Effect of sodium nutrient on chlorophyll a/b rations in C 4 plants. Journal of Plant Physiology, 11,325-332. |
| [19] | Jones HG (1992). Plants and Microclimate. 2nd edn. Cambridge University Press, Cambridge,420-428. |
| [20] | Kenzo T, Ichie T, Ninomiya I, Koike T (2003). Photosynthetic activity in seed wings of Dipterocarpaceae in a masting year: does wing photosynthesis contribute to reproduction? Photosynthetica, 41,551-557. |
| [21] |
Kenzo T, Ichie T, Yoneda R, Kitahashi Y, Watanabe Y, Ninomiya I, Koike T (2004). Interspecific variation of photosynthesis and leaf characteristics in canopy trees of five species of Dipterocarpaceae in a tropical rain forest. Tree Physiology, 24,1187-1192.
DOI URL PMID |
| [22] |
Koch GW, Jeffrey SA, Goulden ML (1994). Diurnal patterns of leaf photosynthesis, conductance and water potential at the top of a lowland rain forest canopy in Cameroon: measurements from the Radeau des Cimes. Tree Physiology, 14,347-360.
URL PMID |
| [23] | Krall JP, Edwards GE (1992). Relationship between photosystem II activity and CO 2 fixation in leaves. Physiologia Plantarum, 86,180-187. |
| [24] | Leakey ADB, Press MC, Scholes JD (2003). Patterns of dynamic irradiance affect the photosynthetic capacity and growth of dipterocarp tree seedlings. Oecologia, 135,184-193. |
| [25] |
Lee DW, Oberbauer SF, Krishnapilay B, Mansor M, Mohamad H, Yap SK (1997). Effects of irradiance and spectral quality on seedling development of two Southeast Asian Hopea species. Oecologia, 110,1-9.
URL PMID |
| [26] |
Ludlow MM, Bjørkman O (1984). Paraheliotropic leaf movement in Siratro as a protective mechanism against drought-induced damaged to primary photosynthesis reactions damage by excessive light and heat. Planta, 161,505-518.
URL PMID |
| [27] | Muraoka H, Tang Y, Terashima I, Koizumi H, Washitani I (2000). Contributions of diffusional limitation, photoinhibition and photorespiration to midday depression of photosynthesis in Arisaema heterophyllum in natural high light. Plant, Cell and Environment, 23,235-250. |
| [28] |
Oberbauer SF, Strain BR, Riechers GH (1987). Field water relations of a wet -tropical forest tree species, Pentaclethra macroloba (Mimosaceae). Oecologia, 71,369-374.
URL PMID |
| [29] | Rascher U, Liebig M, Lüttge U (2000). Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer on site in the field. Plant, Cell and Environment, 23,1397-1405. |
| [30] | Roberts J, Cabral OMR, de Aguiar LF (1990). Stomatal and boundary-layer conductance in an Amazonian terra firm rain forest. Journal of Applied Ecology, 27,336-353. |
| [31] | Scholes JD, Press MC, Zipperlen SW (1997). Differences in light energy utilization and dissipation between dipterocarp rain forest tree seedlings. Oecologia, 109,41-48. |
| [32] | Zhu H (朱华) (2000). Ecology and Biogeography of the Tropical Dipterocarp Rain Forest in Xishuangbanna (西双版纳龙脑香热带雨林生态学与生物地理学研究(第一版))1st edn. Yunnan Science and Technology Press, Kunming,5-18. (in Chinese with English abstract) |
| [1] | 李伟斌, 张红霞, 张玉书, 陈妮娜. 昼夜不对称增温对长白山阔叶红松林碳汇能力的影响[J]. 植物生态学报, 2023, 47(9): 1225-1233. |
| [2] | 蒋海港, 曾云鸿, 唐华欣, 刘伟, 李杰林, 何国华, 秦海燕, 王丽超, 姚银安. 三种藓类植物固碳耗水节律调节作用[J]. 植物生态学报, 2023, 47(7): 988-997. |
| [3] | 任培鑫, 李鹏, 彭长辉, 周晓路, 杨铭霞. 洞庭湖流域植被光合物候的时空变化及其对气候变化的响应[J]. 植物生态学报, 2023, 47(3): 319-330. |
| [4] | 师生波, 周党卫, 李天才, 德科加, 杲秀珍, 马家麟, 孙涛, 王方琳. 青藏高原高山嵩草光合功能对模拟夜间低温的响应[J]. 植物生态学报, 2023, 47(3): 361-373. |
| [5] | 刘海燕, 臧纱纱, 张春霞, 左进城, 阮祚禧, 吴红艳. 磷饥饿下硅藻光系统II光化学反应及其对高光强的响应[J]. 植物生态学报, 2023, 47(12): 1718-1727. |
| [6] | 师生波, 师瑞, 周党卫, 张雯. 低温对高山嵩草叶片光化学和非光化学能量耗散特征的影响[J]. 植物生态学报, 2023, 47(10): 1441-1452. |
| [7] | 薛金儒, 吕肖良. 黄土高原生态工程实施下基于日光诱导叶绿素荧光的植被恢复生产力效益评价[J]. 植物生态学报, 2022, 46(10): 1289-1304. |
| [8] | 吴霖升, 张永光, 章钊颖, 张小康, 吴云飞. 日光诱导叶绿素荧光遥感及其在陆地生态系统监测中的应用[J]. 植物生态学报, 2022, 46(10): 1167-1199. |
| [9] | 靳川, 李鑫豪, 蒋燕, 徐铭泽, 田赟, 刘鹏, 贾昕, 查天山. 黑沙蒿光合能量分配组分在生长季的相对变化与调控机制[J]. 植物生态学报, 2021, 45(8): 870-879. |
| [10] | 武洪敏, 双升普, 张金燕, 寸竹, 孟珍贵, 李龙根, 沙本才, 陈军文. 短期生长环境光强骤增导致典型阴生植物三七光系统受损的机制[J]. 植物生态学报, 2021, 45(4): 404-419. |
| [11] | 周稳, 迟永刚, 周蕾. 基于日光诱导叶绿素荧光的北半球森林物候研究[J]. 植物生态学报, 2021, 45(4): 345-354. |
| [12] | 叶子飘, 于冯, 安婷, 王复标, 康华靖. 植物气孔导度对CO2响应模型的构建[J]. 植物生态学报, 2021, 45(4): 420-428. |
| [13] | 丁键浠, 周蕾, 王永琳, 庄杰, 陈集景, 周稳, 赵宁, 宋珺, 迟永刚. 叶绿素荧光主动与被动联合观测应用前景[J]. 植物生态学报, 2021, 45(2): 105-118. |
| [14] | 李景, 王欣, 王振华, 王斌, 王成章, 邓美凤, 刘玲莉. 臭氧和气溶胶复合污染对杨树叶片光合作用的影响[J]. 植物生态学报, 2020, 44(8): 854-863. |
| [15] | 郭庆华, 胡天宇, 马勤, 徐可心, 杨秋丽, 孙千惠, 李玉美, 苏艳军. 新一代遥感技术助力生态系统生态学研究[J]. 植物生态学报, 2020, 44(4): 418-435. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19