
植物生态学报 ›› 2017, Vol. 41 ›› Issue (6): 610-621.DOI: 10.17521/cjpe.2016.0329
所属专题: 青藏高原植物生态学:植物-土壤-微生物
何为1,2, 杨雪英3, 肖娟4, 张子良1,2, 蒋铮1,2, 袁远爽1,2, 王东1,2, 刘庆1, 尹华军1,4,*( )
)
收稿日期:2017-04-05
接受日期:2016-10-21
出版日期:2017-06-10
发布日期:2017-07-19
通讯作者:
尹华军
作者简介:* 通信作者Author for correspondence (E-mail:基金资助:
Wei HE1,2, Xue-Ying YANG3, Juan XIAO4, Zi-Liang ZHANG1,2, Zheng JIANG1,2, Yuan-Shuang YUAN1,2, Dong WANG1,2, Qing LIU1, Hua-Jun YIN1,4,*()
Received:2017-04-05
Accepted:2016-10-21
Online:2017-06-10
Published:2017-07-19
Contact:
Hua-Jun YIN
About author:KANG Jing-yao(1991-), E-mail: 摘要:
为探究高寒灌丛生态系统根系分泌物碳(C)输入通量对大气氮(N)沉降的响应规律, 该文以青藏高原东缘窄叶鲜卑花(Sibiraea angustata)灌丛为研究对象, 采用根系分泌物野外原位收集法, 分析了不同施N水平(对照N0 = 0 g·m-2·a-1; 低N处理N5 = 5 g·m-2·a-1; 高N处理N10 = 10 g·m-2·a-1)对根系分泌物C输入速率与通量季节动态变化规律的影响。结果表明: (1)窄叶鲜卑花灌丛单位根生物量、单位根长、单位根表面积根系分泌物C输入速率均表现出明显的季节性动态变化, 具体表现为8月> 6月> 10月, 并呈现出与5 cm土壤温度相一致的变化趋势。(2)施N降低了窄叶鲜卑花灌丛单位根生物量、单位根长和单位根表面积根系分泌物C输入速率, 但仅N10处理与对照(N0处理)间存在显著差异(p < 0.05)。(3) N5和N10处理下, 窄叶鲜卑花灌丛细根生物量与N0处理相比分别降低了23.36%和33.84%。(4)由于施N导致根系分泌物C输入速率与细根生物量二者均显著降低, 使得施N对窄叶鲜卑花灌丛根系分泌物C输入通量(g·m-2·a-1)有显著的抑制作用, 并随着施N浓度的增加抑制作用增大。推测其可能的原因是N素富集在一定程度上缓和了植物根系对养分的微生物驱动需求, 从而降低了植物根系分泌物C输入通量, 即N素富集条件下植物采取了低N收益-低C投入的生理策略。该研究结果对于进一步认知不同环境变化下高寒灌丛生态系统根系分泌物C输入及其介导的土壤生物C-养分循环过程具有重要的理论意义。
何为, 杨雪英, 肖娟, 张子良, 蒋铮, 袁远爽, 王东, 刘庆, 尹华军. 氮素富集对青藏高原东缘窄叶鲜卑花灌丛根系分泌物碳输入的影响. 植物生态学报, 2017, 41(6): 610-621. DOI: 10.17521/cjpe.2016.0329
Wei HE, Xue-Ying YANG, Juan XIAO, Zi-Liang ZHANG, Zheng JIANG, Yuan-Shuang YUAN, Dong WANG, Qing LIU, Hua-Jun YIN. Effects of nitrogen enrichment on root exudative carbon inputs in Sibiraea angustata shrubbery at the eastern fringe of Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology, 2017, 41(6): 610-621. DOI: 10.17521/cjpe.2016.0329
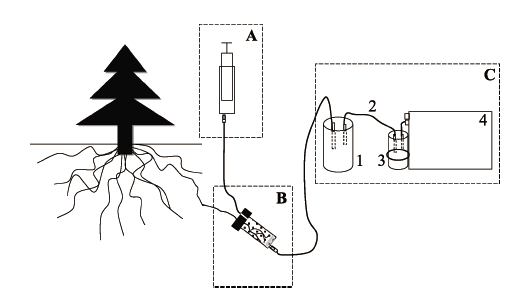
图1 根系分泌物取样装置结构示意图。A, 进样单元。B, 收集单元。C, 抽滤单元。1, 抽滤瓶; 2, 硅胶管; 3, 干燥管; 4, 真空泵。
Fig. 1 Schematic drawing of root exudate collection device. A, Sample injection unit. B, Collection unit. C, Suction filter unit. 1, suction flask; 2, silicone tube; 3, drying tube; 4, vacuum pump.
| 植物类型 Plant type | 文献来源 Reference source | 植物种 Plant species | 单位根生物量根系 分泌物C输入速率 Root exudative C input rate per root biomass (μg·g-1·h-1) | 单位根长根系 分泌物C输入速率 Root exudative C input rate per root length (μg·cm-1·h-1) | 单位根表面积根系 分泌物C输入速率 Root exudative C input rate per root surface area (μg·cm-2·h-1) |
|---|---|---|---|---|---|
| 草本 Herbs | Personeni et al., 2007 | 玉米 Zea mays | - | - | 2.10 |
| Li & Chen, 2011 | 美人蕉 Canna indica | 175.42 | - | - | |
| 风车草 Cyperus flabelliformis | 220.00 | - | - | ||
| 水鬼蕉 Hymenocallis littoralis | 16.79 | - | - | ||
| Cheng at al., 2016 | 小麦 Triticum aestivum | 4.07-1.19 | 0.056-0.012 | ||
| 平均值 Mean | 83.50 | 0.03 | 2.10 | ||
| 乔木 Trees | Phillips et al., 2009 | 火炬松 Pinus taeda | 2.00-40.00 | - | - |
| Phillips et al., 2011 | 火炬松 Pinus taeda | 8.00-17.00 | 0.10-6.00 | - | |
| Meier et al., 2013 | 火炬松 Pinus taeda | 32.00 | - | - | |
| Yin et al., 2013a | 云杉 Picea asperata | - | 0.70 | 9.30 | |
| 冷杉 Abies faxoniana | - | 0.50 | 4.90 | ||
| Yin et al., 2013b | 云杉 Picea asperata | 663.91-414.94 | 0.67-0.39 | 7.99-5.53 | |
| Yin et al., 2014 | 北美鹅掌楸 Liriodendron tulipifera 糖枫 Acer saccharum | 9.58 | - | - | |
| 北美红栎 Quercus rubra 美洲水青冈木 Fagus grandifolia | 17.50 | - | - | ||
| Zhang et al., 2016 | 云杉 Picea asperata | 396.76 | - | - | |
| Li at al., 2014a | 9年生云杉人工林 9-year-old Picea asperata | 441.86 | 3.20 | 2.75 | |
| 13年生云杉人工林 13-year-old Picea asperata | 284.88 | 3.66 | 1.01 | ||
| 31年生云杉人工林 31-year-old Picea asperata | 315.89 | 3.56 | 1.52 | ||
| Xiao, 2013 | 云杉幼苗 Picea asperata seedlings | 79.16 | 0.70 | 9.30 | |
| 冷杉幼苗 Abies faxoniana seedlings | 65.55 | 0.50 | 4.90 | ||
| Qiao, 2015 | 云杉 Picea asperata | 570.93 | 0.50 | 4.44 | |
| 冷杉 Abies faxoniana | 579.71 | 0.51 | 5.52 | ||
| Xiong et al., 2015 | 杉木 Cunninghamia lanceolata | 173.68-138.04 | 0.37-0.14 | 1.67-0.46 | |
| 米槠 Castanopsis carlesi | 92.72-56.16 | 0.34-0.06 | 1.27-0.24 | ||
| 平均值 Mean | 203.07 | 1.29 | 4.05 | ||
| 灌木 Shrubs | 本研究 This study | 窄叶鲜卑花 Sibiraea angustata | 26.22 | 0.02 | 0.29 |
表1 不同研究结果植物根系分泌碳(C)输入速率比较与分析
Table 1 Comparison of root exudative carbon (C) input rates among studies
| 植物类型 Plant type | 文献来源 Reference source | 植物种 Plant species | 单位根生物量根系 分泌物C输入速率 Root exudative C input rate per root biomass (μg·g-1·h-1) | 单位根长根系 分泌物C输入速率 Root exudative C input rate per root length (μg·cm-1·h-1) | 单位根表面积根系 分泌物C输入速率 Root exudative C input rate per root surface area (μg·cm-2·h-1) |
|---|---|---|---|---|---|
| 草本 Herbs | Personeni et al., 2007 | 玉米 Zea mays | - | - | 2.10 |
| Li & Chen, 2011 | 美人蕉 Canna indica | 175.42 | - | - | |
| 风车草 Cyperus flabelliformis | 220.00 | - | - | ||
| 水鬼蕉 Hymenocallis littoralis | 16.79 | - | - | ||
| Cheng at al., 2016 | 小麦 Triticum aestivum | 4.07-1.19 | 0.056-0.012 | ||
| 平均值 Mean | 83.50 | 0.03 | 2.10 | ||
| 乔木 Trees | Phillips et al., 2009 | 火炬松 Pinus taeda | 2.00-40.00 | - | - |
| Phillips et al., 2011 | 火炬松 Pinus taeda | 8.00-17.00 | 0.10-6.00 | - | |
| Meier et al., 2013 | 火炬松 Pinus taeda | 32.00 | - | - | |
| Yin et al., 2013a | 云杉 Picea asperata | - | 0.70 | 9.30 | |
| 冷杉 Abies faxoniana | - | 0.50 | 4.90 | ||
| Yin et al., 2013b | 云杉 Picea asperata | 663.91-414.94 | 0.67-0.39 | 7.99-5.53 | |
| Yin et al., 2014 | 北美鹅掌楸 Liriodendron tulipifera 糖枫 Acer saccharum | 9.58 | - | - | |
| 北美红栎 Quercus rubra 美洲水青冈木 Fagus grandifolia | 17.50 | - | - | ||
| Zhang et al., 2016 | 云杉 Picea asperata | 396.76 | - | - | |
| Li at al., 2014a | 9年生云杉人工林 9-year-old Picea asperata | 441.86 | 3.20 | 2.75 | |
| 13年生云杉人工林 13-year-old Picea asperata | 284.88 | 3.66 | 1.01 | ||
| 31年生云杉人工林 31-year-old Picea asperata | 315.89 | 3.56 | 1.52 | ||
| Xiao, 2013 | 云杉幼苗 Picea asperata seedlings | 79.16 | 0.70 | 9.30 | |
| 冷杉幼苗 Abies faxoniana seedlings | 65.55 | 0.50 | 4.90 | ||
| Qiao, 2015 | 云杉 Picea asperata | 570.93 | 0.50 | 4.44 | |
| 冷杉 Abies faxoniana | 579.71 | 0.51 | 5.52 | ||
| Xiong et al., 2015 | 杉木 Cunninghamia lanceolata | 173.68-138.04 | 0.37-0.14 | 1.67-0.46 | |
| 米槠 Castanopsis carlesi | 92.72-56.16 | 0.34-0.06 | 1.27-0.24 | ||
| 平均值 Mean | 203.07 | 1.29 | 4.05 | ||
| 灌木 Shrubs | 本研究 This study | 窄叶鲜卑花 Sibiraea angustata | 26.22 | 0.02 | 0.29 |
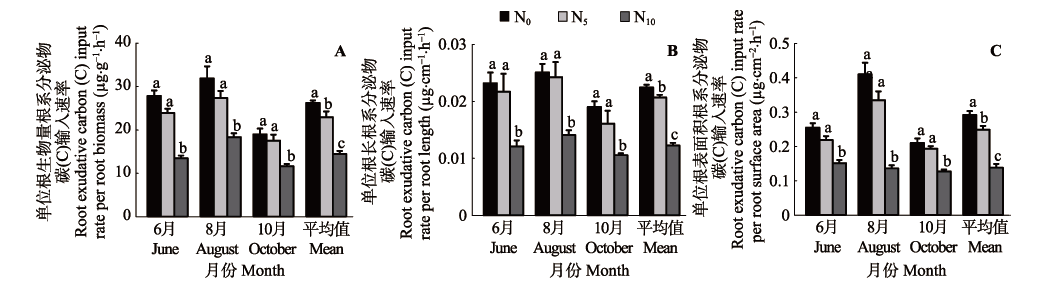
图2 不同施氮(N)处理下窄叶鲜卑花灌丛根系分泌物碳(C)输入速率差异(平均值±标准偏差, n = 3)。A, 单位根生物量根系分泌物C输入速率。B, 单位根长根系分泌物C输入速率。C, 单位根表面积根系分泌物C输入速率。不同小写字母表示同一采样时间各处理间在p < 0.05水平上差异显著。N0、N5和N10为3种施氮梯度(N0: 0 g·m-2·a-1; N5: 5 g·m-2·a-1; N10: 10 g·m-2·a-1)。
Fig. 2 Differences in root exudative carbon (C) input rates in Sibiraea angustata shrubbery among different nitrogen fertilization treatments (mean ± SD, n = 3). A, Root exudative C input rate per root biomass. B, Root exudative C input rate per root length. C, Root exudative C input rate per root surface area. Different lowercase letters indicate significant differences (p < 0.05) among treatments on a given sampling date. Three nitrogen addition levels: N0 (0 g·m-2·a-1), N5 (5 g·m-2·a-1), N10 (10 g·m-2·a-1).
| 施N N fertilization | 采样日期 Sampling date | 采样日期×施N Sampling date × N fertilization | |
|---|---|---|---|
| 单位根生物量根系分泌物C输入速率 Root exudative C input rates per root biomass | <0.001 | <0.001 | 0.255 |
| 单位根长根系分泌物C输入速率 Root exudative C input rates per root length | <0.001 | 0.019 | 0.844 |
| 单位根表面积根系分泌物C输入速率 Root exudative C input rates per root surface area | <0.001 | <0.001 | <0.001 |
表2 窄叶鲜卑花灌丛根系分泌物碳(C)输入速率对施氮(N)和采样日期的重复测量方差分析p值
Table 2 Summary of repeated measures ANOVA showing the p values for responses of root exudative carbon (C) input rates of Sibiraea angustata shrubbery to nitrogen (N) fertilization and sampling date
| 施N N fertilization | 采样日期 Sampling date | 采样日期×施N Sampling date × N fertilization | |
|---|---|---|---|
| 单位根生物量根系分泌物C输入速率 Root exudative C input rates per root biomass | <0.001 | <0.001 | 0.255 |
| 单位根长根系分泌物C输入速率 Root exudative C input rates per root length | <0.001 | 0.019 | 0.844 |
| 单位根表面积根系分泌物C输入速率 Root exudative C input rates per root surface area | <0.001 | <0.001 | <0.001 |
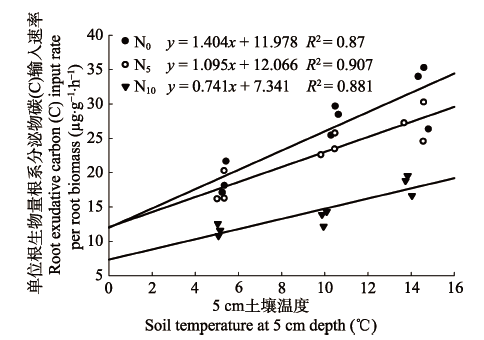
图3 窄叶鲜卑花灌丛单位根生物量根系分泌物碳(C)输入速率与5 cm土壤温度的相关性分析。N0、N5和N10为3种施氮梯度(N0: 0 g·m-2·a-1; N5: 5 g·m-2·a-1; N10: 10 g·m-2·a-1)。
Fig. 3 Correlation analysis between root exudative carbon (C) input rate per root biomass and soil temperature at 5 cm depth. Three nitrogen addition levels: N0 (0 g·m-2·a-1), N5 (5 g·m-2·a-1), N10 (10 g·m-2·a-1).
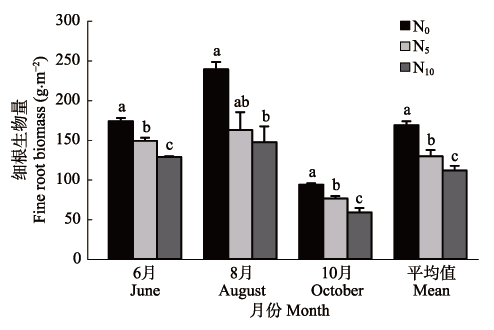
图4 不同施N处理下窄叶鲜卑花灌丛细根生物量变化(平均值±标准偏差, n = 3)。不同小写字母表示同一采样时间各处理间在p < 0.05水平上差异显著。N0、N5和N10为3种施氮梯度(N0: 0 g·m-2·a-1; N5: 5 g·m-2·a-1; N10: 10 g·m-2·a-1)。
Fig. 4 Changes in fine root biomass in Sibiraea angustata shrubbery under different nitrogen fertilization treatments (mean ± SD, n = 3). Different lowercase letters indicate significant differences among treatments on a given sampling date (p < 0.05). Three nitrogen addition levels: N0 (0 g·m-2·a-1), N5 (5 g·m-2·a-1), N10 (10 g·m-2·a-1).
| 处理 Treatment | 单位根生物量根系分泌物C输入速率 Root exudative C input rate per root biomass (mg·g-1·d-1) | 细根生物量 Fine root biomass (g·m-2) | 根系分泌物C通量 C flux of root exudates (g·m-2·a-1) |
|---|---|---|---|
| N0 | 0.68 ± 0.02a | 169.17 ± 47.83a | 16.89 ± 0.31a |
| N5 | 0.57 ± 0.04b | 129.65 ± 81.71b | 9.58 ± 0.24b |
| N10 | 0.36 ± 0.02c | 111.93 ± 58.49c | 5.81 ± 0.12c |
表3 不同施氮(N)处理下窄叶鲜卑花灌丛根系分泌物碳(C)通量(平均值±标准偏差, n = 3)
Table 3 Root exudative carbon (C) flux in Sibiraea angustata shrubbery under different nitrogen (N) fertilization treatments (mean ± SD, n = 3)
| 处理 Treatment | 单位根生物量根系分泌物C输入速率 Root exudative C input rate per root biomass (mg·g-1·d-1) | 细根生物量 Fine root biomass (g·m-2) | 根系分泌物C通量 C flux of root exudates (g·m-2·a-1) |
|---|---|---|---|
| N0 | 0.68 ± 0.02a | 169.17 ± 47.83a | 16.89 ± 0.31a |
| N5 | 0.57 ± 0.04b | 129.65 ± 81.71b | 9.58 ± 0.24b |
| N10 | 0.36 ± 0.02c | 111.93 ± 58.49c | 5.81 ± 0.12c |
| 处理 Treatment | 2015-06 | 2015-08 | 2015-10 | ||||
|---|---|---|---|---|---|---|---|
| NO3--N (mg·kg-1) | NH4+-N (mg·kg-1) | NO3--N (mg·kg-1) | NH4+-N (mg·kg-1) | NO3--N (mg·kg-1) | NH4+-N (mg·kg-1) | ||
| N0 | 20.88 ± 0.91a | 7.44 ± 0.16a | 23.92 ± 0.23a | 8.66 ± 0.54a | 25.88 ± 0.83a | 7.63 ± 0.28a | |
| N5 | 22.19 ± 0.71a | 12.57 ± 0.64b | 24.51 ± 0.31a | 13.21 ± 0.27b | 26.21 ± 0.24a | 12.42 ± 0.59b | |
| N10 | 30.38 ± 0.71b | 14.29 ± 0.85b | 28.98 ± 0.65b | 16.18 ± 0.56b | 29.38 ± 0.17b | 13.94 ± 0.71b | |
表4 不同施N水平下土壤NO3--N、NH4+-N含量(平均值±标准偏差, n =3)
Table 4 Changes in nitrate nitrogen and ammonium nitrogen under different nitrogen fertilization treatments (mean ± SD, n = 3)
| 处理 Treatment | 2015-06 | 2015-08 | 2015-10 | ||||
|---|---|---|---|---|---|---|---|
| NO3--N (mg·kg-1) | NH4+-N (mg·kg-1) | NO3--N (mg·kg-1) | NH4+-N (mg·kg-1) | NO3--N (mg·kg-1) | NH4+-N (mg·kg-1) | ||
| N0 | 20.88 ± 0.91a | 7.44 ± 0.16a | 23.92 ± 0.23a | 8.66 ± 0.54a | 25.88 ± 0.83a | 7.63 ± 0.28a | |
| N5 | 22.19 ± 0.71a | 12.57 ± 0.64b | 24.51 ± 0.31a | 13.21 ± 0.27b | 26.21 ± 0.24a | 12.42 ± 0.59b | |
| N10 | 30.38 ± 0.71b | 14.29 ± 0.85b | 28.98 ± 0.65b | 16.18 ± 0.56b | 29.38 ± 0.17b | 13.94 ± 0.71b | |
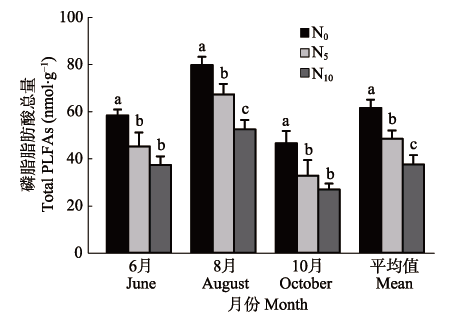
图5 不同施N处理下土壤微生物磷脂脂肪酸(PLFAs)总量变化(平均值±标准偏差, n = 3)。不同小写字母表示同一采样时间各处理间在p < 0.05水平上差异显著。N0、N5和N10为3种施氮梯度(N0: 0 g·m-2·a-1; N5: 5 g·m-2·a-1; N10: 10 g·m-2·a-1)。
Fig. 5 Changes in total soil microbial phospholipid fatty acid (PLFAs) under different nitrogen fertilization treatments (mean ± SD, n = 3). Different lowercase letters indicate significant differences (p < 0.05) among treatments on a given sampling date. Three nitrogen addition levels: N0 (0 g·m-2·a-1), N5 (5 g·m-2·a-1), N10 (10 g·m-2·a-1).
| [1] | Björk RG, Majdi H, Klemedtsson L, Lewisjonsson L, Molau U (2007). Long-term warming effects on root morphology, root mass distribution, and microbial activity in two dry tundra plant communities in northern Sweden.New Phytologist, 176, 862-673. |
| [2] | Cheng LS, Cheng W, Bian ZF, Lei SG (2016). Wheat root exudation and soil nitrogen transformations in farmland reclaimed from coal gangue.Journal of Ecology and Rural Environment, 32, 397-403. (in Chinese with English abstract)[ 程林森, 程伟, 卞正富, 雷少刚 (2016). 煤矸石复垦对小麦根系分泌特征和土壤氮转化的影响. 生态与农村环境学报, 32, 397-403.] |
| [3] | Cheng WX, Kuzyakov Y, Zobel RW, Wright S (2005). Root effects on soil organic matter decomposition. In: Zobel RW, Wright SF eds. Roots and Soil Management: Interactions Between Roots and the Soil, Agronomy Monograph No. 48.American Society of Agronomy, Madison, USA. 119-143. |
| [4] | Dijkstra FA, Cheng WX (2007). Interactions between soil and tree roots accelerate long-term soil carbon decomposition.Ecology Letters, 10, 1046-1053. |
| [5] | Drake JE, Darby BA, Giasson MA, Kramer MA, Phillips RP, Finzi AC (2013). Stoichiometry constrains microbial response to root exudation-insights from a model and a field experiment in a temperate forest.Biogeosciences, 10, 821-838. |
| [6] | Fenn ME, Poth MA, Terry JD, Blubaugh TJ (2005). Nitrogen mineralization and nitrification in a mixed-conifer forest in southern California: Controlling factors, fluxes, and nitrogen fertilization response at a high and low nitrogen deposition site.Canadian Journal of Forest Research-revue Canadian de Recherche Forestiere, 35, 1464-1486. |
| [7] | Fleischer K, Rebel KT, Molen MK, Erisman JW, Wassen MJ, Loon EE, Montagnani L, Gough CM, Herbst M, Janssens IA, Gianelle D, Dolman AJ (2013). The contribution of nitrogen deposition to the photosynthetic capacity of forests. Global Biogeochemical Cycles, 27, 187-199. |
| [8] | Galloway JN, Townsend AR, Erisman JW, Bekunda MA, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008). Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions.Science, 320, 889-892. |
| [9] | Gao Q, Yang XC, Yin CY, Liu Q (2014). Estimation of biomass allocation and carbon density in alpine dwarf shrubs in Garzê Zangzu Autonomous Prefecture of Sichuan Province, China.Chinese Journal of Plant Ecology, 38, 355-365. (in Chinese with English abstract)[ 高巧, 阳小成, 尹春英, 刘庆 (2014). 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算. 植物生态学报, 38, 355-365.] |
| [10] | He HL, Yang XC, Wang D, Sun YY, Yin CY, Li T, Li YX, Zhou GY, Zhang L, Liu Q (2015). Ecological stoichiometric characteristics of soil carbon, nitrogen and phosphorus of Sibiraea angustata shrub in eastern Qinghai-Tibetan Plateau. Chinese Journal of Applied Environmental Biology, 21, 1128-1135. (in Chinese with English abstract)[ 贺合亮, 阳小成, 王东, 孙誉育, 尹春英, 李婷, 黎云祥, 周国英, 张林, 刘庆 (2015). 青藏高原东部窄叶鲜卑花灌丛土壤C、N、P生态化学计量学特征. 应用与环境生物学报, 21, 1128-1135.] |
| [11] | He JS, Wang ZQ, Fang JY (2004). The below-ground ecology under global change: Problems and prospect.Chinese Science Bulletin, 49, 1226-1233. (in Chinese)[ 贺金生, 王政权, 方精云 (2004). 全球变化下的地下生态学: 问题与展望. 科学通报, 49, 1226-1233.] |
| [12] | Hodge A, Grayson SJ, Ord BG (1996). A novel method for characterization and quantification of plant root exudates.Plant and Soil, 184, 97-104. |
| [13] | Inagaki M, Inagaki Y, Kamo K, Titin J (2009). Fine-root production in response to nutrient application at three forest plantations in sabah, malaysia: Higher nitrogen and phosphorus demand byAcacia mangium. Journal of Forest Research, 14, 178-182. |
| [14] | Jacqueline A, Peterson A, Kalbitz K (2005). Short-term response on the quantity and quality of rhizo-deposited carbon from Norway spruce exposed to low and high N inputs.Journal of Plant Nutrition and Soil Science, 168, 687-693. |
| [15] | Jin M, Li Y, Wang SL, Zhang XL, Lei L (2012). Alpine shrubs biomass and its distribution characteristics in Qilian Mountains.Arid Land Geography, 35, 952-959. (in Chinese with English abstract)[ 金铭, 李毅, 王顺利, 张学龙, 雷蕾 (2012). 祁连山高山灌丛生物量及其分配特征. 干旱区地理, 35, 952-959.] |
| [16] | Jones DL, Hodge A, Kuzyakov Y (2004). Plant and mycorrhizal regulation of rhizodeposition.New Phytologist, 163, 459-480. |
| [17] | Joslin JD, Wolfe MH, Hanson PJ (2001). Factors controlling the timing of root elongation intensity in a mature upland oak stand.Plant and Soil, 228, 201-212. |
| [18] | Kopacek J, Cosby BJ, Evans CD, Hruska JD, Moldan F, Oulehle F, Santrůckova H, Tahovska k, Wright RF (2013). Nitrogen, organic carbon and sulphur cycling in terrestrial ecosystems: Linking nitrogen saturation to carbon limitation of soil microbial processes. Biogeochemistry, 115, 33-51. |
| [19] | Kuzyakov Y, Cheng WX (2001). Photosynthesis controls of rhizosphere respiration and organic matter decomposition.Soil Biology & Biochemistry, 33, 1915-1925. |
| [20] | LeBauer DS, Treseder KK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.Ecology, 89, 371-379. |
| [21] | Lei L, Liu XD, Wang SL, Li Y, Zhang XL (2011). Assignment rule of alpine shrubs biomass and its relationships to environmental factors in Qilian Mountains.Ecology and Environmental Sciences, 20, 1602-1607. (in Chinese with English abstract)[ 雷蕾, 刘贤德, 王顺利, 李毅, 张学龙 (2011). 祁连山高山灌丛生物量分配规律及其与环境因子的关系. 生态环境学报, 20, 1602-1607.] |
| [22] | Li DJ, Mo JM, FANG YT, Peng SL, Gundersen P (2003). Impact of nitrogen deposition on forest plants.Acta Ecologica Sinica, 23, 1891-1900. (in Chinese with English abstract)[ 李德军, 莫江明, 方运霆, 彭少麟, Per Gundersen (2003). 氮沉降对森林植物的影响. 生态学报, 23, 1891-1900.] |
| [23] | Li HY, Chen ZH (2011). Growth and dissolved organic carbon exudates from roots of three wetland plants.Journal of Tropical and Subtropical Botany, 6, 536-542. (in Chinese with English abstract)[ 李海燕, 陈章和 (2011). 三种湿地植物的生长及根系溶解性有机碳分泌物研究. 热带亚热带植物学报, 6, 536-542.] |
| [24] | Li J, Jiang XM, Yin HJ, Yin CY, Wei YH, Liu Q (2014a). Root exudates and soil microbes in three Picea asperata plantations with different stand ages.Chinese Journal of Applied Ecology, 25, 325-332. (in Chinese with English abstract)[ 李娇, 蒋先敏, 尹华军, 尹春英, 魏宇航, 刘庆 (2014a). 不同林龄云杉人工林的根系分泌物与土壤微生物. 应用生态学报, 25, 325-332.] |
| [25] | Li J, Yin CY, Zhou XB, Wei YH, Gao Q, Liu Q (2014b). Effects of nitrogen addition on soil respiration ofSibiraea angustata shrub in the eastern margin of Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 34, 5558-5569. (in Chinese with English abstract)[ 李娇, 尹春英, 周晓波, 魏宇航, 高巧, 刘庆 (2014b). 施氮对青藏高原东缘窄叶鲜卑花灌丛土壤呼吸的影响. 生态学报, 34, 5558-5569.] |
| [26] | Li X, Duan ZQ (2013). Progress on the research methods for root exudates.Genomics and Applied Biology, 32, 540-547. (in Chinese with English abstract)[ 李讯, 段增强 (2013). 植物根系分泌物的研究方法. 基因组学与应用生物学, 32, 540-547.] |
| [27] | Liu ZX, Zhu TH, Zhang J (2005). Research advances in root exudates and rhizosphere microorganisms of forest trees.World Forestry Research, 18(6), 25-31. (in Chinese with English abstract)[ 刘子雄, 朱天辉, 张建 (2005). 林木根系分泌物与根际微生物研究进展. 世界林业研究, 18(6), 25-31.] |
| [28] | Lu CQ, Tian HQ (2007). Spatial and temporal patterns of nitrogen deposition in China synthesis of observational data. Journal of Geophysical Research, 112(D22), 1-10. |
| [29] | Luo YQ, Zhao XY, Li MX (2012). Ecological effect of plant root exudates and related affecting factors: A review.Chinese Journal of Applied Ecology, 23, 3496-3504. (in Chinese with English abstract)[ 罗永清, 赵学勇, 李美霞 (2012). 植物根系分泌物生态效应及其影响因素研究综述. 应用生态学报, 23, 3496-3504.] |
| [30] | Magill AH, Aber JD, Currie WS, Nadelhoffer KJ, Martin ME, McDowell WH, Melillo JM, Steudler P (2004). Ecosystems response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA.Forest Ecology and Management, 196, 7-28. |
| [31] | Majdi H (2001). Changes in fine root production and longevity in relation to water and nutrient availability in a Norwaye spruce stand in northern Sweden.Tree Physiology, 21, 1057-1061. |
| [32] | Meier IC, Avis PG, Phillips RP (2013). Fungal communities influence root exudation rates in pine seedlings.FEMS Microbiology Ecology, 83, 585-595. |
| [33] | Nadelhoffer KJ (2000). The potential effects of nitrogen deposition on fine root production in forest ecosystems.New Phytologist, 147, 131-139. |
| [34] | Paterson E, Gebbing T, Abel C, Sim A, Telfer G (2007). Rhizodeposition shapes rhizosphere microbial community structure in organic soil.New Phytologist, 173, 600-610. |
| [35] | Personeni E, Nguyen C, Marcha P, Pages L (2007). Experimental evaluation of an efflux-influx model of C exudation by individual apical root segments.Journal of Experimental Botany, 58, 2091-2099. |
| [36] | Phillips RP, Bernhardt ES, Schlesinger WH (2009). Elevated CO2 increases root exudation from loblolly pine (Pinus taeda) seedlings as an N-mediated response. Tree Physiology, 29, 1513-1523. |
| [37] | Phillips RP, Fahey TJ (2007). Fertilization effects on fine root biomass, rhizosphere microbes and respiratory fluxes in hard wood forest soils.New Phytologist, 176, 655-664. |
| [38] | Phillips RP, Finzi AC, Bernhardt ES (2011). Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation.Ecology Letters, 14, 187-194. |
| [39] | Qiao MF (2015). Effects of Night Warming and Nitrogen Fertilization on Root Exudates of the Seedlings of Two Subalpine Coniferous Species in Western Sichuan. PhD dissertation, Sichuan University, Chengdu. (in Chinese with English abstract)[ 乔明锋 (2015). 增温条件下川西亚高山针叶林云杉和冷杉根系分泌物及其生态学效应研究. 博士学位论文, 四川大学, 成都.] |
| [40] | Talhelm AF, Burton AJ, Pregitzer KS, Campione MA (2013). Chronic nitrogen deposition reduces the abundance of dominant forest understory and groundcover species.Forest Ecology and Management, 293, 39-48. |
| [41] | Tu SX, Wu J (2010). A review on research methods of root exudates.Ecology and Environmental Sciences, 19, 2493-2500. (in Chinese with English abstract)[ 涂书新, 吴佳 (2010). 植物根系分泌物研究方法评述. 生态环境学报, 19, 2493-2500.] |
| [42] | Uselman SM, Qualls RG, Thomas RB (1999). A test of a potential short cut in the nitrogen cycle: The role of exudation of symbiotically fixed nitrogen from the roots of a N-fixing tree and the effects of increased atmospheric CO2 and temperature.Plant and Soil, 210, 21-32. |
| [43] | Volder A, Gifford RM, Evans JR (2007). Effects of elevated atmospheric CO2, cutting frequency, and differential day/night atmospheric warming on root growth and turnover of phalaris swards.Global Change Biology, 13, 1040-1052. |
| [44] | Wells CE, Glenn DM, Eissenstat D (2005). Soil insects alter fine root demography in peach (Prunus persica). Plant, Cell & Environment, 25, 431-439. |
| [45] | White DC, Stair JO, Ringelberg DB (1996). Quantitative comparisons of in situ microbial biodiversity by signature biomarker analysis. Journal of Industrial Microbiology, 17, 185-196. |
| [46] | Wu N (1998). The community types and biomass of Sibiraea angustata scrub and their relationship with environmental factors in northwestern Sichuan. Acta Botanica Sinica, 40, 860-870. (in Chinese with English abstract)[ 吴宁 (1998). 川西北窄叶鲜卑花灌丛的类型和生物量及其与环境因子的关系. 植物学报, 40, 860-870.] |
| [47] | Xia J, Wan S (2013). Independent effects of warming and nitrogen addition on plant phenology in the Inner Mongolian steppe.Annals of Botany, 111, 1207-1217. |
| [48] | Xiao J (2013). Effects of Night Warming and Nitrogen Fertilization on Root Exudates of the Seedlings of Two Subalpine Coniferous Species in Western Sichuan. PhD dissertation, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu. (in Chinese with English abstract)[ 肖娟 (2013). 夜间增温和施N对两种川西亚高山针叶树幼苗根系分泌物的影响研究. 博士学位论文, 中国科学院成都生物研究所, 成都.] |
| [49] | Xiong DC, Huang JX, Chen GS, Xie JS, Yang YS (2015). A preliminary report on the fine root exudate ofCastanopsis carlesii and Cunninghamia lanceolata in subtropical zone. Journal of Subtropical Resources and Environment, 10(1), 83-86. (in Chinese)[ 熊德成, 黄锦学, 陈光水, 谢锦升, 杨玉盛 (2015). 中亚热带米槠和杉木细根分泌物研究初报. 亚热带资源与环境学报, 10(1), 83-86.] |
| [50] | Yang JH (2015). Influence of Nitrogen Deposition and Rainfall Manipulation on Concentration of Forest Tree Root Exudates. Master degree dissertation, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang. (in Chinese with English abstract)[ 杨建华 (2015). N沉降与降水变化对森林根系分泌物的影响. 硕士学位论文, 中国科学院沈阳应用生态研究所, 沈阳.] |
| [51] | Yin HJ, Li Y, Xiao J, Xu ZF, Cheng XY, Liu Q (2013a). Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming.Global Change Biology, 19, 2158-2167. |
| [52] | Yin HJ, Wheeler E, Phillips RP (2014). Root-induced changes in nutrient cycling in forests depend on exudation rates. Soil Biology & Biochemistry, 78, 213-221. |
| [53] | Yin HJ, Xiao J, Li YF, Chen Z, Cheng XY, Zhao CZ, Liu Q (2013b). Warming effects on root morphological and physiological traits the potential consequences on soil C dynamics as altered root exudation.Agricultural and Forest Meteorology, 180, 287-296. |
| [54] | Zeng ZY, Liu QJ, Zhang JP, Zeng HQ, Cai Z (2005). A study on the pertinence of measure factors and organic biomass of shrub.Acta Agriculturae Universitatis Jiangxiensis, 27, 694-699. (in Chinese with English abstract)[ 曾珍英, 刘琪璟, 张建萍, 曾慧卿, 蔡哲 (2005). 灌木各测树因子相关性以及器官生物量相关性的研究. 江西农业大学学报, 27, 694-699.] |
| [55] | Zhang YC, Hou SG, Pang HX (2012). Preliminary study on spatiotemporal pattern of climate change over Tibet Plateau during past millennium.Marine Geology & Quaternary Geology, 32(3), 135-146. (in Chinese with English abstract)[ 张彦成, 侯书贵, 庞洪喜 (2012). 青藏高原地区近千年气候变化的时空特征. 海洋地质与第四纪地质, 32(3), 135-146.] |
| [56] | Zhang ZL, Qiao MF, Li DD, Yin HJ, Liu Q (2016). Do warming-induced changes in quantity and stoichiometry of root exudation promote soil N transformations via stimulation of soil nitrifiers, denitrifiers and ammonifiers?European Journal of Soil Biology, 74, 60-68. |
| [57] | Zheng XH, Fu CB, Xu XK, Yan XD, Huang Y, Han SH, Hu F, Chen GX (2002). The Asian nitrogen cycle case study.AMBIO, 31, 79-87. |
| [58] | Zong N, Shi PL, Jiang J, Xiong DP, Meng FS, Song MH, Zhang XZ, Shen ZX (2013). Interactive effects of short-term nitrogen enrichment and simulated grazing on ecosystem respiration in an alpine meadow on the Tibetan Plateau.Acta Ecologica Sinica, 33, 6191-6201. (in Chinese with English abstract)[ 宗宁, 石培礼, 蒋婧, 熊定鹏, 孟丰收, 宋明华, 张宪洲, 沈振西 (2013). 短期N素添加和模拟放牧对青藏高原高寒草甸生态系统呼吸的影响. 生态学报, 33, 6191-6201.] |
| [1] | 黄玲, 王榛, 马泽, 杨发林, 李岚, SEREKPAYEV Nurlan, NOGAYEV Adilbek, 侯扶江. 长期放牧和氮添加对黄土高原典型草原长芒草种群生长的影响[J]. 植物生态学报, 2024, 48(3): 317-330. |
| [2] | 颜辰亦, 龚吉蕊, 张斯琦, 张魏圆, 董学德, 胡宇霞, 杨贵森. 氮添加对内蒙古温带草原土壤活性有机碳的影响[J]. 植物生态学报, 2024, 48(2): 229-241. |
| [3] | 耿雪琪, 唐亚坤, 王丽娜, 邓旭, 张泽凌, 周莹. 氮添加增加中国陆生植物生物量并降低其氮利用效率[J]. 植物生态学报, 2024, 48(2): 147-157. |
| [4] | 舒韦维, 杨坤, 马俊旭, 闵惠琳, 陈琳, 刘士玲, 黄日逸, 明安刚, 明财道, 田祖为. 氮添加对红锥不同序级细根形态和化学性状的影响[J]. 植物生态学报, 2024, 48(1): 103-112. |
| [5] | 赵艳超, 陈立同. 土壤养分对青藏高原高寒草地生物量响应增温的调节作用[J]. 植物生态学报, 2023, 47(8): 1071-1081. |
| [6] | 苏炜, 陈平, 吴婷, 刘岳, 宋雨婷, 刘旭军, 刘菊秀. 氮添加与干季延长对降香黄檀幼苗非结构性碳水化合物、养分与生物量的影响[J]. 植物生态学报, 2023, 47(8): 1094-1104. |
| [7] | 李红琴, 张法伟, 仪律北. 高寒草甸表层土壤和优势植物叶片的化学计量特征对降水改变和氮添加的响应[J]. 植物生态学报, 2023, 47(7): 922-931. |
| [8] | 何斐, 李川, Faisal SHAH, 卢谢敏, 王莹, 王梦, 阮佳, 魏梦琳, 马星光, 王卓, 姜浩. 丛枝菌根菌丝桥介导刺槐-魔芋间碳转运和磷吸收[J]. 植物生态学报, 2023, 47(6): 782-791. |
| [9] | 张雅琪, 庞丹波, 陈林, 曹萌豪, 何文强, 李学斌. 荒漠草原土壤氨氧化细菌群落结构对氮添加和枯落物输入的响应[J]. 植物生态学报, 2023, 47(5): 699-712. |
| [10] | 罗来聪, 赖晓琴, 白健, 李爱新, 方海富, Nasir SHAD, 唐明, 胡冬南, 张令. 氮添加背景下土壤真菌和细菌对不同种源入侵植物乌桕生长特征的影响[J]. 植物生态学报, 2023, 47(2): 206-215. |
| [11] | 安凡, 李宝银, 钟全林, 程栋梁, 徐朝斌, 邹宇星, 张雪, 邓兴宇, 林秋燕. 不同种源刨花楠苗木生长与主要功能性状对氮添加的响应[J]. 植物生态学报, 2023, 47(12): 1693-1707. |
| [12] | 葛萍, 李昂, 王银柳, 姜良超, 牛国祥, 哈斯木其尔, 王彦兵, 薛建国, 赵威, 黄建辉. 草甸草原温室气体排放对氮添加量的非线性响应[J]. 植物生态学报, 2023, 47(11): 1483-1492. |
| [13] | 董六文, 任正炜, 张蕊, 谢晨笛, 周小龙. 功能多样性比物种多样性更好解释氮添加对高寒草地生物量的影响[J]. 植物生态学报, 2022, 46(8): 871-881. |
| [14] | 谢欢, 张秋芳, 陈廷廷, 曾泉鑫, 周嘉聪, 吴玥, 林惠瑛, 刘苑苑, 尹云锋, 陈岳民. 氮添加促进丛枝菌根真菌和根系协作维持土壤磷有效性[J]. 植物生态学报, 2022, 46(7): 811-822. |
| [15] | 马炬峰, 辛敏, 徐陈超, 祝琬莹, 毛传澡, 陈欣, 程磊. 丛枝菌根真菌与氮添加对不同根形态基因型水稻氮吸收的影响[J]. 植物生态学报, 2021, 45(7): 728-737. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19