
植物生态学报 ›› 2016, Vol. 40 ›› Issue (9): 893-901.DOI: 10.17521/cjpe.2016.0163
所属专题: 青藏高原植物生态学:群落生态学
岳楷1, 杨万勤1,2, 彭艳1, 黄春萍1,3, 张川1, 吴福忠1,2,*( )
)
收稿日期:2016-05-09
接受日期:2016-07-23
出版日期:2016-09-10
发布日期:2016-09-29
通讯作者:
吴福忠
基金资助:
Kai YUE1, Wan-Qin YANG1,2, Yan PENG1, Chun-Ping HUANG1,3, Chuan ZHANG1, Fu-Zhong WU1,2,*()
Received:2016-05-09
Accepted:2016-07-23
Online:2016-09-10
Published:2016-09-29
Contact:
Fu-Zhong WU
摘要:
溪流广泛分布于高寒森林地表, 凋落于其中的林木凋落物的分解是整个森林生态系统物质循环的重要环节, 水体流动过程中的冲刷和淋洗作用及其他独特的环境条件可能显著影响凋落物中木质素的降解。该研究采用凋落袋法对比研究了岷江上游高寒森林4种典型且初始质量差异显著的凋落叶, 即康定柳(Salix paraplesia)、高山杜鹃(Rhododendron lapponicum)、方枝柏(Sabina saltuaria)和四川红杉(Larix mastersiana), 在不同生境(林下、溪流和河岸带)下分解过程中木质素残留质量和浓度(质量百分率)的动态变化特征。经过两年的分解, 发现溪流显著促进了凋落叶中木质素的降解; 同一物种凋落叶在不同生境下木质素残留质量差异显著(p < 0.05), 整体表现为溪流<河岸带<林下; 在凋落叶分解的初期木质素有明显的降解, 其浓度表现为先降低后升高, 但不同物种之间存在显著(p < 0.05)的差异; 在整个分解过程中, 木质素残留质量总体呈现出了降低的趋势。此外, 生境类型、分解时期和区域性环境因子(温度、pH值和营养元素的有效性)能显著影响木质素的降解率。这些结果表明, 传统上认为木质素在凋落叶分解初期相对稳定的观点可能并不准确, 其浓度很可能是先下降后升高, 这也与有关木质素动态的最新研究结果相一致。另一方面, 在不同分解时期和不同生境下, 凋落叶木质素降解率表现出了显著差异, 表明区域性环境因子在凋落叶分解和木质素降解过程中具有重要的作用。
岳楷, 杨万勤, 彭艳, 黄春萍, 张川, 吴福忠. 高寒森林溪流对凋落叶分解过程中木质素降解的影响. 植物生态学报, 2016, 40(9): 893-901. DOI: 10.17521/cjpe.2016.0163
Kai YUE, Wan-Qin YANG, Yan PENG, Chun-Ping HUANG, Chuan ZHANG, Fu-Zhong WU. Effects of streams on lignin degradation during foliar litter decomposition in an alpine forest. Chinese Journal of Plant Ecology, 2016, 40(9): 893-901. DOI: 10.17521/cjpe.2016.0163
| 生境 Habitat | AT (℃) | C (g·kg-1) | N (g·kg-1) | P (g·kg-1) | pH | ||
|---|---|---|---|---|---|---|---|
| 林下 Forest floor | 2.0 ± 5.2 | 126 ± 26 | 5.8 ± 1.1 | 1.2 ± 0.2 | 6.6 ± 0.02 | ||
| 生境 Habitat | AT (°C) | HCO3- (mg·L-1) | NH4+ (mg·L-1) | NO3-(mg·L-1) | PO43- (μg·L-1) | pH | FV (m·s-1) |
| 溪流 Stream | 5.1 ± 2.6 | 13.9 ± 1.96 | 0.10 ± 0.05 | 0.29 ± 0.07 | 7.85 ± 0.38 | 6.6 ± 0.4 | 0.53 ± 0.15 |
| 河岸带 Riparian zone | 4.8 ± 3.4 | 19.7 ± 1.33 | 0.04 ± 0.02 | 0.34 ± 0.08 | 7.84 ± 0.41 | 6.9 ± 0.3 | 0.05 ± 0.01 |
表1 不同生境下凋落叶分解过程中环境因子特征(平均值±标准偏差, n = 90)
Table 1 Characteristics of environmental conditions of different habitats during the process of foliar litter decomposition (mean ± SD, n = 90)
| 生境 Habitat | AT (℃) | C (g·kg-1) | N (g·kg-1) | P (g·kg-1) | pH | ||
|---|---|---|---|---|---|---|---|
| 林下 Forest floor | 2.0 ± 5.2 | 126 ± 26 | 5.8 ± 1.1 | 1.2 ± 0.2 | 6.6 ± 0.02 | ||
| 生境 Habitat | AT (°C) | HCO3- (mg·L-1) | NH4+ (mg·L-1) | NO3-(mg·L-1) | PO43- (μg·L-1) | pH | FV (m·s-1) |
| 溪流 Stream | 5.1 ± 2.6 | 13.9 ± 1.96 | 0.10 ± 0.05 | 0.29 ± 0.07 | 7.85 ± 0.38 | 6.6 ± 0.4 | 0.53 ± 0.15 |
| 河岸带 Riparian zone | 4.8 ± 3.4 | 19.7 ± 1.33 | 0.04 ± 0.02 | 0.34 ± 0.08 | 7.84 ± 0.41 | 6.9 ± 0.3 | 0.05 ± 0.01 |
| 物种 Species | C (%) | N (%) | P (%) | 木质素 Lignin (%) | C:N | C:P | N:P | Lignin:N |
|---|---|---|---|---|---|---|---|---|
| 康定柳 S. paraplesia | 34.8 ± 0.9c | 2.64 ± 0.15a | 0.17 ± 0.01a | 24.7 ± 1.3d | 13.2 ± 0.8d | 207 ± 19.7c | 15.7 ± 1.7a | 9.38 ± 0.8c |
| 高山杜鹃 R. lapponicum | 38.6 ± 1.1b | 0.69 ± 0.10d | 0.10 ± 0.02d | 29.8 ± 0.8b | 57.2 ± 10.2a | 375 ± 53.6a | 6.75 ± 1.5c | 44.3 ± 8.3a |
| 方枝柏 S. saltuaria | 46.9 ± 1.8a | 1.05 ± 0.06c | 0.15 ± 0.01b | 28.1 ± 0.8c | 45.1 ± 3.9b | 304 ± 12.6b | 6.79 ± 0.7c | 26.9 ± 1.8b |
| 四川红杉 L. mastersiana | 37.5 ± 0.5b | 1.59 ± 0.11b | 0.12 ± 0.01c | 37.8 ± 1.0a | 23.6 ± 1.8c | 320 ± 24.6b | 13.6 ± 0.8b | 30.1 ± 2.1b |
表2 康定柳、高山杜鹃、方枝柏和四川红杉凋落叶初始质量特征(平均数±标准偏差, n = 9)
Table 2 Initial chemical properties of Salix paraplesia, Rhododendron lapponicum, Sabina saltuaria, and Larix mastersiana foliar litters (mean ± SD, n = 9)
| 物种 Species | C (%) | N (%) | P (%) | 木质素 Lignin (%) | C:N | C:P | N:P | Lignin:N |
|---|---|---|---|---|---|---|---|---|
| 康定柳 S. paraplesia | 34.8 ± 0.9c | 2.64 ± 0.15a | 0.17 ± 0.01a | 24.7 ± 1.3d | 13.2 ± 0.8d | 207 ± 19.7c | 15.7 ± 1.7a | 9.38 ± 0.8c |
| 高山杜鹃 R. lapponicum | 38.6 ± 1.1b | 0.69 ± 0.10d | 0.10 ± 0.02d | 29.8 ± 0.8b | 57.2 ± 10.2a | 375 ± 53.6a | 6.75 ± 1.5c | 44.3 ± 8.3a |
| 方枝柏 S. saltuaria | 46.9 ± 1.8a | 1.05 ± 0.06c | 0.15 ± 0.01b | 28.1 ± 0.8c | 45.1 ± 3.9b | 304 ± 12.6b | 6.79 ± 0.7c | 26.9 ± 1.8b |
| 四川红杉 L. mastersiana | 37.5 ± 0.5b | 1.59 ± 0.11b | 0.12 ± 0.01c | 37.8 ± 1.0a | 23.6 ± 1.8c | 320 ± 24.6b | 13.6 ± 0.8b | 30.1 ± 2.1b |
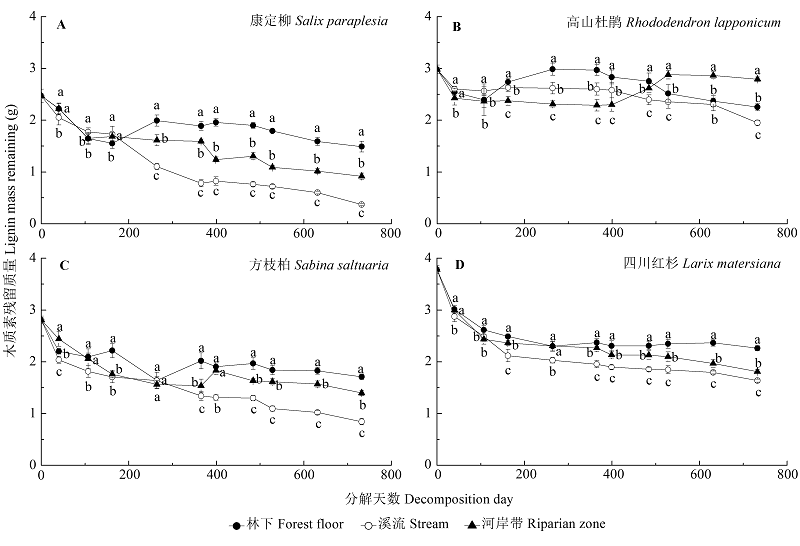
图1 不同生境下康定柳(A)、高山杜鹃(B)、方枝柏(C)、四川红杉(D)凋落叶分解过程中木质素残留质量(g)的动态特征(平均值±标准偏差, n = 9)。不同小写字母表示同一分解时期同一物种在不同生境下木质素残留质量差异显著(p < 0.05)。
Fig. 1 Dynamics of lignin mass remaining (g) in the decomposing foliar litter of Salix paraplesia (A), Rhododendron lapponicum (B), Sabina saltuaria (C), and Larix mastersiana (D) under different habitat conditions (mean ± SD, n = 9). Different lowercase letters indicate significant (p < 0.05) differences of lignin mass remaining for a given litter species in a specific decomposition period under different habitat conditions.
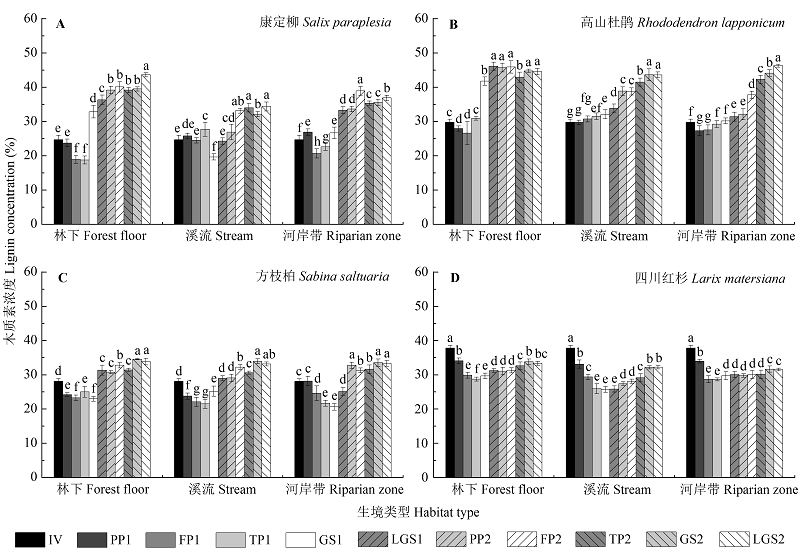
图2 不同生境下康定柳(A)、高山杜鹃(B)、方枝柏(C)、四川红杉(D)凋落叶分解过程中木质素浓度(质量百分率, %)的动态特征(平均值±标准偏差, n = 9)。不同小写字母表示同一物种凋落叶在同一生境的不同分解时期木质素浓度差异显著(p < 0.05)。FP, 冻结期; GS, 生长季节; IV, 初始值; LGS, 生长季节后期; PP, 冻结初期; TP, 融化期; 1, 第一年; 2, 第二年。
Fig. 2 Dynamics of lignin concentration (percent litter mass, %) during Salix paraplesia (A), Rhododendron lapponicum (B), Sabina saltuaria (C), and Larix mastersiana (D) foliar litter decomposition (p < 0.05) under different habitat conditions (mean ± SD, n = 9). Different lowercase letters indicate significant (p < 0.05) differences of lignin concentration among different decomposition periods for a given litter species incubated in a specific type of habitat. FP, freezing period; GS, growing season; IV, initial value; LGS, late growing season; PP, pre-freezing period; TP, thawing period; 1, first year; 2, second year.
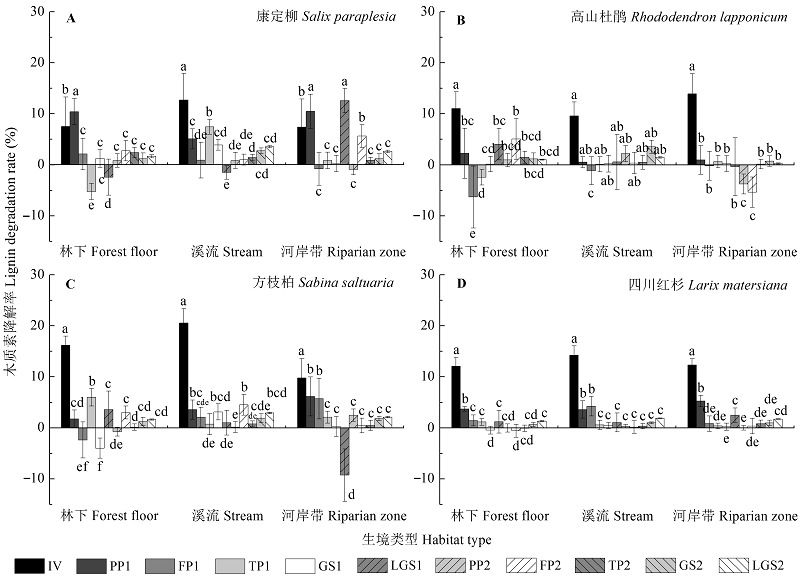
图3 不同生境下康定柳(A)、高山杜鹃(B)、方枝柏(C)、四川红杉(D)凋落叶分解过程中木质素降解率(%/月)的动态特征(平均值±标准偏差, n = 9)。不同小写字母表示同一物种凋落叶在同一生境下不同分解时期木质素降解率差异显著(p < 0.05)。FP, 冻结期; IV, 初始值; GS, 生长季节; LGS, 生长季节后期; PP, 冻结初期; TP, 融化期; 1, 第一年; 2, 第二年。
Fig. 3 Dynamics of lignin degradation rate (%/month) during Salix paraplesia (A), Rhododendron lapponicum (B), Sabina saltuaria (C), and Larix mastersiana (D) foliar litter decomposition (p < 0.05) under different habitat conditions (mean ± SD, n = 9). Different lowercase letters indicate significant (p < 0.05) differences of lignin degradation rate among different decomposition periods for a given litter species incubated in a specific type of habitat. FP, freezing period; GS, growing season; IV, initial value; LGS, late growing season; PP, pre-freezing period; TP, thawing period; 1, first year; 2, second year.
| 影响因子 Influence factor | 自由度 Degree of freedom | F | p |
|---|---|---|---|
| 物种 Species | 3 | 165.753 | < 0.001 |
| 生境 Habitat | 2 | 75.197 | < 0.001 |
| 时期 Period | 9 | 504.141 | < 0.001 |
| 物种×生境 Species × habitat | 6 | 40.353 | < 0.001 |
| 物种×时期 Species × period | 27 | 17.003 | < 0.001 |
| 生境×时期 Habitat × period | 18 | 18.317 | < 0.001 |
| 物种×生境×时期 Species × habitat × period | 54 | 12.020 | < 0.001 |
表3 凋落叶分解过程中木质素降解率与物种、生境和分解时期的重复测量方差分析
Table 3 Repeated-measure ANOVA analysis on the effects of litter species, habitat type, and decomposition period on lignin degradation rate during litter decomposition process
| 影响因子 Influence factor | 自由度 Degree of freedom | F | p |
|---|---|---|---|
| 物种 Species | 3 | 165.753 | < 0.001 |
| 生境 Habitat | 2 | 75.197 | < 0.001 |
| 时期 Period | 9 | 504.141 | < 0.001 |
| 物种×生境 Species × habitat | 6 | 40.353 | < 0.001 |
| 物种×时期 Species × period | 27 | 17.003 | < 0.001 |
| 生境×时期 Habitat × period | 18 | 18.317 | < 0.001 |
| 物种×生境×时期 Species × habitat × period | 54 | 12.020 | < 0.001 |
| 生境 Habitat | 回归式 Regression model | ||||
|---|---|---|---|---|---|
| a0 | a1X1 | a2X2 | a3X3 | a4X4 | |
| 林下 Forest floor | ŷ = 0.424 | -0.017 C:N (0.353) | +0.042 C (0.607) | ||
| 溪流 Stream | ŷ = 1.602 | -0.031 Lignin:N (0.785) | +13.231 P (0.874) | ||
| 河岸带 Riparian zone | ŷ = -7.311 | +0.032 Lignin (0.783) | +20.108 P (0.888) | +0.164 N:P (0.932) | +0.090 C (0.940) |
表4 凋落叶木质素两年总降解率(%/月)与初始质量的逐步回归分析
Table 4 Stepwise regression analysis between lignin degradation rate (%/month) of the 2 years and foliar litter initial chemical properties
| 生境 Habitat | 回归式 Regression model | ||||
|---|---|---|---|---|---|
| a0 | a1X1 | a2X2 | a3X3 | a4X4 | |
| 林下 Forest floor | ŷ = 0.424 | -0.017 C:N (0.353) | +0.042 C (0.607) | ||
| 溪流 Stream | ŷ = 1.602 | -0.031 Lignin:N (0.785) | +13.231 P (0.874) | ||
| 河岸带 Riparian zone | ŷ = -7.311 | +0.032 Lignin (0.783) | +20.108 P (0.888) | +0.164 N:P (0.932) | +0.090 C (0.940) |
| 林下 Forest floor | AT | C | N | P | pH | ||
|---|---|---|---|---|---|---|---|
| 康定柳 Salix paraplesia | 26.925*** | 35.094*** | 1.987 | 0.340 | 6.194* | ||
| 高山杜鹃 Rhododendron lapponicum | 16.022*** | 0.064 | 5.700* | 17.816*** | 2.431 | ||
| 方枝柏 Sabina saltuaria | 10.134** | 1.037 | 23.348*** | 23.681*** | 8.314** | ||
| 四川红杉 Larix mastersiana | 30.336*** | 7.748** | 32.560*** | 10.076** | 13.489*** | ||
| 溪流 Stream | AT | HCO3- | NH4+ | NO3- | PO43- | pH | FV |
| 康定柳 Salix paraplesia | 0.001 | 0.572 | 2.692 | 13.248*** | 0.522 | 6.208* | 0.385 |
| 高山杜鹃 Rhododendron lapponicum | 1.286 | 1.722 | 6.088* | 8.832** | 1.612 | 1.652 | 1.590 |
| 方枝柏 Sabinasaltuaria | 1.245 | 4.809* | 7.579** | 8.964** | 0.001 | 6.454* | 0.103 |
| 四川红杉 Larix mastersiana | 2.815 | 2.179 | 4.681* | 11.866** | 0.063 | 5.594* | 0.053 |
| 河岸带 Riparian zone | AT | HCO3- | NH4+ | NO3- | PO43- | pH | FV |
| 康定柳 Salix paraplesia | 35.148*** | 5.748* | 12.267** | 0.256 | 1.305 | 16.431*** | 2.540 |
| 高山杜鹃 Rhododendron lapponicum | 3.702 | 2.822 | 2.029 | 1.369 | 0.001 | 7.300** | 5.752* |
| 方枝柏 Sabina saltuaria | 1.564 | 4.024* | 6.775* | 4.609* | 0.115 | 3.545 | 6.232* |
| 四川红杉 Larix mastersiana | 36.978*** | 15.189*** | 19.985*** | 4.305* | 0.055 | 0.371 | 11.602** |
表5 不同生境下凋落叶分解过程中木质素降解率(%/月)与环境因子回归分析的F值
Table 5 F-value for the regression analysis between lignin degradation rate (%/month) and environmental factors under different habitats during foliar litter decomposition
| 林下 Forest floor | AT | C | N | P | pH | ||
|---|---|---|---|---|---|---|---|
| 康定柳 Salix paraplesia | 26.925*** | 35.094*** | 1.987 | 0.340 | 6.194* | ||
| 高山杜鹃 Rhododendron lapponicum | 16.022*** | 0.064 | 5.700* | 17.816*** | 2.431 | ||
| 方枝柏 Sabina saltuaria | 10.134** | 1.037 | 23.348*** | 23.681*** | 8.314** | ||
| 四川红杉 Larix mastersiana | 30.336*** | 7.748** | 32.560*** | 10.076** | 13.489*** | ||
| 溪流 Stream | AT | HCO3- | NH4+ | NO3- | PO43- | pH | FV |
| 康定柳 Salix paraplesia | 0.001 | 0.572 | 2.692 | 13.248*** | 0.522 | 6.208* | 0.385 |
| 高山杜鹃 Rhododendron lapponicum | 1.286 | 1.722 | 6.088* | 8.832** | 1.612 | 1.652 | 1.590 |
| 方枝柏 Sabinasaltuaria | 1.245 | 4.809* | 7.579** | 8.964** | 0.001 | 6.454* | 0.103 |
| 四川红杉 Larix mastersiana | 2.815 | 2.179 | 4.681* | 11.866** | 0.063 | 5.594* | 0.053 |
| 河岸带 Riparian zone | AT | HCO3- | NH4+ | NO3- | PO43- | pH | FV |
| 康定柳 Salix paraplesia | 35.148*** | 5.748* | 12.267** | 0.256 | 1.305 | 16.431*** | 2.540 |
| 高山杜鹃 Rhododendron lapponicum | 3.702 | 2.822 | 2.029 | 1.369 | 0.001 | 7.300** | 5.752* |
| 方枝柏 Sabina saltuaria | 1.564 | 4.024* | 6.775* | 4.609* | 0.115 | 3.545 | 6.232* |
| 四川红杉 Larix mastersiana | 36.978*** | 15.189*** | 19.985*** | 4.305* | 0.055 | 0.371 | 11.602** |
| 1 | Berg B (2014). Decomposition patterns for foliar litter—A theory for influencing factors.Soil Biology & Biochem- istry, 78, 222-232. |
| 2 | Berg B, Kjønaas O, Johansson M-B, Erhagen B, Åkerblom S (2015). Late stage pine litter decomposition: Relationship to litter N, Mn, and acid unhydrolyzable residue (AUR) concentrations and climatic factors.Forest Ecology and Management, 358, 41-47. |
| 3 | Berg B, McClaugherty C (2014). Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. 3rd edn. Springer, Berlin. |
| 4 | Boyero L, Pearson RG, Gessner MO, Barmuta LA, Ferreira V, Graça MAS, Dudgeon D, Boulton AJ, Callisto M, Chauvet E, Helson JE, Bruder A, Albariño RJ, Yule CM, Arunachalam M, Davies JN, Figueroa R, Flecker AS, Ramírez A, Death RG, Iwata T, Mathooko JM, Mathuriau C, Gonçalves JF, Moretti MS, Jinggut T, Lamothe S, M’Erimba C, Ratnarajah L, Schindler MH, Castela J, Buria LM, Cornejo A, Villanueva VD, West DC (2011). A global experiment suggests climate warming will not accelerate litter decomposition in streams but might reduce carbon sequestration.Ecology Letters, 14, 289-294. |
| 5 | Bradford MA, Berg B, Maynard DS, Wieder WR, Wood SA (2016). Understanding the dominant controls on litter decomposition.Journal of Ecology, 104, 229-238. |
| 6 | Bradford MA, Warren II RJ, Baldrian P, Crowther TW, Maynard DS, Oldfield EE, Wieder WR, Wood SA, King JR (2014). Climate fails to predict wood decomposition at regional scales.Nature Climate Change, 4, 625-630. |
| 7 | Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested HM, Santiago LS, Wardle DA, Wright IJ, Aerts R, Allison SD, van Bodegom P, Brovkin V, Chatain A, Callaghan TV, Díaz S, Garnier E, Gurvich DE, Kazakou E, Klein JA, Read J, Reich PB, Soudzilovskaia NA, Vaieretti MV, Westoby M (2008). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide.Ecology Letters, 11, 1065-1071. |
| 8 | Ferreira V, Raposeiro PM, Pereira A, Cruz AM, Costa AC, Graça MAS, Gonçalves V (2016). Leaf litter decomposi- tion in remote oceanic island streams is driven by microbes and depends on litter quality and environmental conditions.Freshwater Biology, 61, 783-799. |
| 9 | García-Palacios P, Prieto I, Ourcival J-M, Hättenschwiler S (2016a). Disentangling the litter quality and soil microbial contribution to leaf and fine root litter decomposition responses to reduced rainfall.Ecosystems, 19, 490-503. |
| 10 | García-Palacios P, Shaw EA, Wall DH, Hättenschwiler S (2016b). Temporal dynamics of biotic and abiotic drivers of litter decomposition.Ecology Letters, 19, 554-563. |
| 11 | Gessner MO, Chauvet E, Dobson M (1999). A perspective on leaf litter breakdown in streams.Oikos, 85, 377-384. |
| 12 | Graça MA, Ferreira V, Canhoto C, Encalada AC, Guerrero- Bolaño F, Wantzen KM, Boyero L (2015). A conceptual model of litter breakdown in low order streams.International Review of Hydrobiology, 100, 1-12. |
| 13 | He W, Wu FZ, Yang WQ, Tan B, Zhao YY, Wu QQ, He M (2016). Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest.Ecosystems, 19, 115-128. |
| 14 | He W, Wu FZ, Yang WQ, Wu QQ, He M, Zhao YY (2013). Effect of snow patches on leaf litter mass loss of two shrubs in an alpine forest.Chinese Journal of Plant Ecology, 37, 306-316. (in Chinese with English abstract)[何伟, 吴福忠, 杨万勤, 武启骞, 何敏, 赵野逸 (2013). 雪被斑块对高山森林两种灌木凋落叶质量损失的影响. 植物生态学报, 37, 306-316.] |
| 15 | Klotzbücher T, Kaiser K, Guggenberger G, Gatzek C, Kalbitz K (2011). A new conceptual model for the fate of lignin in decomposing plant litter.Ecology, 92, 1052-1062. |
| 16 | Li H, Wu FZ, Yang WQ, Xu LY, Ni XY, He J, Tan B, Hu Y (2016). Effects of forest gaps on litter lignin and cellulose dynamics vary seasonally in an alpine forest.Forests, 7, 27. |
| 17 | Martínez A, Larrañaga A, Pérez J, Descals E, Pozo J (2014). Temperature affects leaf litter decomposition in low- order forest streams: Field and microcosm approaches.FEMS Microbiology Ecology, 87, 257-267. |
| 18 | Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Adair EC, Brandt LA, Hart SC (2007). Global-scale similarities in nitrogen release patterns during long-term decomposition.Science, 315, 361-364. |
| 19 | Prescott CE (2005). Do rates of litter decomposition tell us anything we really need to know?Forest Ecology and Management, 220, 66-74. |
| 20 | Wallace JB, Eggert S, Meyer JL, Webster J (1999). Effects of resource limitation on a detrital-based ecosystem.Ecological Monographs, 69, 409-442. |
| 21 | Yue K, Yang WQ, Peng CH, Peng Y, Zhang C, Huang CP, Tan Y, Wu FZ (2016). Foliar litter decomposition in an alpine forest meta-ecosystem on the eastern Tibetan Plateau. Science of the Total Environment, 566-567, 279-287. |
| 22 | Yue K, Yang WQ, Peng Y, Zhang C, Huang CP, Wu FZ (2015a). Carbon, nitrogen and phosphorus dynamics during winter foliar litter decomposition in an alpine forest river in the upper reaches of the Minjiang River.Chinese Journal of Applied and Environmental Biology, 21, 301-307. (in Chinese with English abstract)[岳楷, 杨万勤, 彭艳, 张川, 黄春萍, 吴福忠 (2015a). 岷江上游高山森林冬季河流中凋落叶碳氮和磷元素动态特征. 应用与环境生物学报, 21, 301-307.] |
| 23 | Yue K, Yang WQ, Peng Y, Zhang C, Huang CP, Wu FZ (2015b). Foliar litter mass loss in winter in an alpine forest river in the upper reaches of the Minjiang River.Resources and Environment in the Yangtze Basin, 24, 1177-1184. (in Chinese with English abstract)[岳楷, 杨万勤, 彭艳, 张川, 黄春萍, 吴福忠 (2015b). 岷江上游高山森林凋落叶在冬季河流中的质量损失特征. 长江流域资源与环境, 24, 1177-1184.] |
| 24 | Zhang C, Yang WQ, Yue K, Huang CP, Peng Y, Wu FZ (2015). Soluble nitrogen and soluble phosphorus dynamics during foliar litter decomposition in winter in alpine forest streams.Chinese Journal of Applied Ecology, 26, 1601-1608. (in Chinese with English abstract)[张川, 杨万勤, 岳楷, 黄春萍, 彭艳, 吴福忠 (2015). 高山森林溪流冬季不同时期凋落物分解中水溶性氮和磷的动态特征. 应用生态学报, 26, 1601-1608.] |
| 25 | Zhu JX, He XH, Wu FZ, Yang WQ, Tan B (2012). Decomposi- tion of Abies faxoniana litter varies with freeze-thaw stages and altitudes in subalpine/alpine forests of southwest China.Scandinavian Journal of Forest Research, 27, 586-596. |
| [1] | 李闪闪 刘学勤. 长江中游河漫滩湿地植物功能性状分析[J]. 植物生态学报, 2024, 48(5): 601-611. |
| [2] | 董劭琼, 侯东杰, 曲孝云, 郭柯. 柴达木盆地植物群落样方数据集[J]. 植物生态学报, 2024, 48(4): 534-540. |
| [3] | 盘远方, 潘良浩, 邱思婷, 邱广龙, 苏治南, 史小芳, 范航清. 中国沿海红树林树高变异与环境适应机制[J]. 植物生态学报, 2024, 48(4): 483-495. |
| [4] | 薛志方, 刘彤, 王立生, 宋继虎, 陈宏阳, 徐玲, 袁也. 额尔齐斯河流域主要支流平原河谷林群落结构及特征[J]. 植物生态学报, 2024, 48(3): 390-402. |
| [5] | 牛一迪, 蔡体久. 大兴安岭北部次生林演替过程中物种多样性的变化及其影响因子[J]. 植物生态学报, 2024, 48(3): 349-363. |
| [6] | 陈雨婷, 马松梅, 张丹, 张林, 王春成. 新疆同域分布梭梭和白梭梭多样性格局及其形成机制[J]. 植物生态学报, 2024, 48(1): 56-67. |
| [7] | 肖兰, 董标, 张琳婷, 邓传远, 李霞, 姜德刚, 林勇明. 渤海无居民海岛主要植被类型群落特征[J]. 植物生态学报, 2024, 48(1): 127-134. |
| [8] | 王雨婷, 刘旭婧, 唐驰飞, 陈玮钰, 王美娟, 向松竹, 刘梅, 杨林森, 傅强, 晏召贵, 孟红杰. 神农架极小种群植物庙台槭群落特征及种群动态[J]. 植物生态学报, 2024, 48(1): 80-91. |
| [9] | 陈保冬, 付伟, 伍松林, 朱永官. 菌根真菌在陆地生态系统碳循环中的作用[J]. 植物生态学报, 2024, 48(1): 1-20. |
| [10] | 袁雅妮, 周哲, 陈彬洲, 郭垚鑫, 岳明. 基于功能性状的锐齿槲栎林共存树种生态策略差异[J]. 植物生态学报, 2023, 47(9): 1270-1277. |
| [11] | 李娜, 唐士明, 郭建英, 田茹, 王姗, 胡冰, 罗永红, 徐柱文. 放牧对内蒙古草地植物群落特征影响的meta分析[J]. 植物生态学报, 2023, 47(9): 1256-1269. |
| [12] | 任悦, 高广磊, 丁国栋, 张英, 赵珮杉, 柳叶. 不同生长期樟子松外生菌根真菌群落物种组成及其驱动因素[J]. 植物生态学报, 2023, 47(9): 1298-1309. |
| [13] | 李安艳, 黄先飞, 田源斌, 董继兴, 郑菲菲, 夏品华. 贵州草海草-藻型稳态转换过程中叶绿素a的变化及其影响因子[J]. 植物生态学报, 2023, 47(8): 1171-1181. |
| [14] | 杨鑫, 任明迅. 环南海区域红树物种多样性分布格局及其形成机制[J]. 植物生态学报, 2023, 47(8): 1105-1115. |
| [15] | 于笑, 纪若璇, 任天梦, 夏新莉, 尹伟伦, 刘超. 中国北方蒙古莸群落的分布、特征和分类[J]. 植物生态学报, 2023, 47(8): 1182-1192. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19