
植物生态学报 ›› 2016, Vol. 40 ›› Issue (12): 1257-1266.DOI: 10.17521/cjpe.2016.0218
所属专题: 生态化学计量
周正虎, 王传宽*
出版日期:2016-12-10
发布日期:2016-12-30
通讯作者:
王传宽
基金资助:Zheng-Hu ZHOU, Chuan-Kuan WANG*
Online:2016-12-10
Published:2016-12-30
Contact:
Chuan-Kuan WANG
摘要:
土壤碳(C)、氮(N)、磷(P)化学计量特征会显著影响微生物的生长、群落结构、生物量C:N:P化学计量及其代谢活动。然而生态系统演替过程中土壤-微生物C:N:P化学计量的时间格局及其协调关系还不明确。为此, 该研究收集了2016年5月以前发表的文献中19个生态系统演替序列(包括13个森林、6个草地生态系统)的土壤-微生物生物量C:N:P研究结果, 整合分析了其中土壤-微生物生态化学计量的时间动态, 结果表明: (1)生态系统演替过程中土壤C:N没有一致的时间格局, 而土壤C:P和N:P均随演替进程显著增加, 其中土壤C:N:P与演替时间之间线性关系的斜率与相应演替序列的初始土壤有机C含量呈负相关关系。(2)演替进程中土壤-微生物生物量C:N:P没有一致的时间格局。(3)微生物生物量C占土壤有机C百分比(qMBC)、微生物生物量N占土壤全N百分比、微生物生物量P占土壤全P百分比均随着演替进程而显著增加, 即单位资源所能支持的微生物生物量随着演替进程而增加, 这与宏观生态系统演替理论相符。(4) qMBC随着土壤C:N、C:P和N:P以及C:N、C:P和N:P化学计量不平衡性(即土壤C:N、C:P和N:P分别除以微生物生物量C:N、C:P和N:P)的增加而减小; 其中, C:N、C:P和N:P化学计量不平衡性解释了qMBC变异性的37%-57%, 是演替时间解释率的7-17倍, 表明土壤-微生物生态化学计量关系对qMBC演替动态有重要影响。该研究强调了生态化学计量学理论和生态系统演替理论在土壤微生物时间动态研究中的重要作用, 表明适当地融合生态学宏观理论于土壤微生物研究可以加深对土壤-微生物生态过程的认识。
周正虎, 王传宽. 生态系统演替过程中土壤与微生物碳氮磷化学计量关系的变化. 植物生态学报, 2016, 40(12): 1257-1266. DOI: 10.17521/cjpe.2016.0218
Zheng-Hu ZHOU, Chuan-Kuan WANG. Changes of the relationships between soil and microbes in carbon, nitrogen and phosphorus stoichiometry during ecosystem succession. Chinese Journal of Plant Ecology, 2016, 40(12): 1257-1266. DOI: 10.17521/cjpe.2016.0218
| 参考文献 Reference | 生态系统类型 Ecosystem type | 年龄阶段数 No. of age stages | 演替年龄跨度 Span of successional age (a) | Csoil | Nsoil | Psoil | Cmic | Nmic | Pmic |
|---|---|---|---|---|---|---|---|---|---|
| An et al., 2009 | 草地 Grassland | 8 | 78 | √ | √ | NA | √ | √ | NA |
| Banning et al., 2008 | 森林 Forest | 7 | 27 | √ | √ | NA | √ | √ | NA |
| 森林 Forest | 7 | 27 | √ | √ | NA | √ | √ | NA | |
| Cao et al., 2008 | 森林 Forest | 4 | 24 | √ | √ | NA | √ | √ | NA |
| Hu et al., 2016 | 森林 Forest | 5 | 13 | √ | √ | NA | √ | √ | NA |
| Jia et al., 2005 | 森林 Forest | 5 | 36 | √ | √ | NA | √ | √ | NA |
| Jia, 2006 | 草地 Grassland | 9 | 27 | √ | √ | √ | √ | √ | √ |
| Jiang et al., 2007 | 草地 Grassland | 6 | 24 | √ | √ | NA | √ | √ | NA |
| Jiang et al., 2009 | 草地 Grassland | 6 | 139 | √ | √ | NA | √ | √ | NA |
| Liu et al., 2012 | 森林 Forest | 5 | 39 | √ | √ | NA | √ | NA | √ |
| Liu et al., 2010 | 森林 Forest | 7 | 101 | √ | √ | NA | √ | √ | √ |
| Liu et al., 2013 | 森林 Forest | 5 | 56 | √ | √ | √ | √ | √ | NA |
| Singh et al., 2001 | 森林 Forest | 5 | 58 | √ | √ | √ | √ | √ | √ |
| Xiao et al., 2013 | 草地 Grassland | 8 | 30 | √ | √ | NA | √ | √ | NA |
| Xue et al., 2008 | 森林 Forest | 6 | 31 | √ | √ | √ | √ | √ | √ |
| Xue et al., 2007 | 森林 Forest | 9 | 51 | √ | √ | √ | √ | √ | √ |
| Xue et al., 2009 | 草地 Grassland | 11 | 51 | √ | √ | √ | √ | √ | √ |
| Yang et al., 2014 | 森林 Forest | 4 | 49 | √ | √ | √ | √ | √ | √ |
| Zhu et al., 2012 | 森林 Forest | 4 | 51 | √ | √ | NA | √ | √ | NA |
表1 满足本研究筛选要求的数据介绍
Table 1 Summary of the data sets compiled in this study
| 参考文献 Reference | 生态系统类型 Ecosystem type | 年龄阶段数 No. of age stages | 演替年龄跨度 Span of successional age (a) | Csoil | Nsoil | Psoil | Cmic | Nmic | Pmic |
|---|---|---|---|---|---|---|---|---|---|
| An et al., 2009 | 草地 Grassland | 8 | 78 | √ | √ | NA | √ | √ | NA |
| Banning et al., 2008 | 森林 Forest | 7 | 27 | √ | √ | NA | √ | √ | NA |
| 森林 Forest | 7 | 27 | √ | √ | NA | √ | √ | NA | |
| Cao et al., 2008 | 森林 Forest | 4 | 24 | √ | √ | NA | √ | √ | NA |
| Hu et al., 2016 | 森林 Forest | 5 | 13 | √ | √ | NA | √ | √ | NA |
| Jia et al., 2005 | 森林 Forest | 5 | 36 | √ | √ | NA | √ | √ | NA |
| Jia, 2006 | 草地 Grassland | 9 | 27 | √ | √ | √ | √ | √ | √ |
| Jiang et al., 2007 | 草地 Grassland | 6 | 24 | √ | √ | NA | √ | √ | NA |
| Jiang et al., 2009 | 草地 Grassland | 6 | 139 | √ | √ | NA | √ | √ | NA |
| Liu et al., 2012 | 森林 Forest | 5 | 39 | √ | √ | NA | √ | NA | √ |
| Liu et al., 2010 | 森林 Forest | 7 | 101 | √ | √ | NA | √ | √ | √ |
| Liu et al., 2013 | 森林 Forest | 5 | 56 | √ | √ | √ | √ | √ | NA |
| Singh et al., 2001 | 森林 Forest | 5 | 58 | √ | √ | √ | √ | √ | √ |
| Xiao et al., 2013 | 草地 Grassland | 8 | 30 | √ | √ | NA | √ | √ | NA |
| Xue et al., 2008 | 森林 Forest | 6 | 31 | √ | √ | √ | √ | √ | √ |
| Xue et al., 2007 | 森林 Forest | 9 | 51 | √ | √ | √ | √ | √ | √ |
| Xue et al., 2009 | 草地 Grassland | 11 | 51 | √ | √ | √ | √ | √ | √ |
| Yang et al., 2014 | 森林 Forest | 4 | 49 | √ | √ | √ | √ | √ | √ |
| Zhu et al., 2012 | 森林 Forest | 4 | 51 | √ | √ | NA | √ | √ | NA |
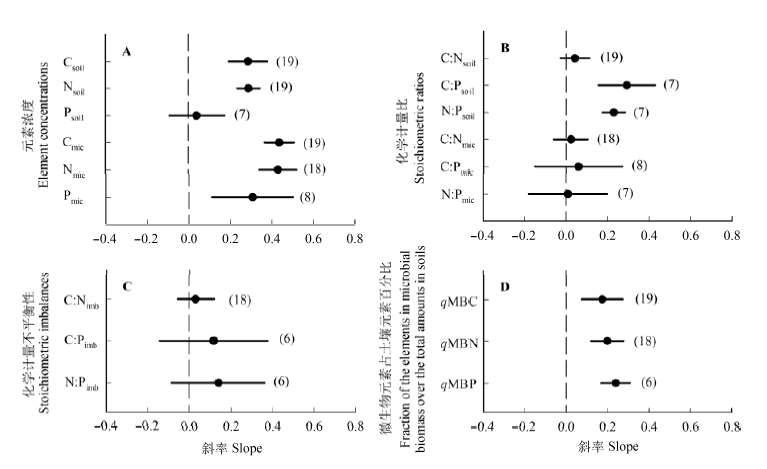
图1 不同演替序列土壤和微生物C、N、P的浓度(A)、化学计量比(B)、化学计量不平衡性(C), 以及微生物生物量元素占土壤元素百分比(D)与演替时间线性关系斜率的平均值和95%置信区间。图中点和误差线分别表示斜率的平均值和95%置信区间。括号内数字表示报道该变量的演替序列个数。Csoil, 土壤有机C; Nsoil, 土壤全N; Psoil, 土壤全P; Cmic, 微生物生物量C; Nmic, 微生物生物量N; Pmic, 微生物生物量P; C:Nsoil, 土壤C:N; C:Psoil, 土壤C:P; N:Psoil, 土壤N:P; C:Nmic, 微生物生物量C:N; C:Pmic, 微生物生物量C:P; N:Pmic, 微生物生物量N:P; C:Nimb, C:N化学计量不平衡性; C:Pimb, C:P化学计量不平衡性; N:Pimb, N:P化学计量不平衡性; qMBC, 微生物生物量C占土壤有机C百分比; qMBN, 微生物生物量N占土壤全N百分比; qMBP, 微生物生物量P占土壤全P百分比。
Fig. 1 Means and 95% confidence interval of the slopes of the linear relationships between soil and microbial C, N, P concentrations (A), stoichiometric ratios (B), stoichiometric imbalances (C), fraction of the elements in microbial biomass over the total amounts in soils (D) and the successional age. The dots and error bars represent the means and 95% confidence intervals, respectively; values in brackets are the numbers of age sequences. Csoil, soil organic C; Nsoil, soil total N; Psoil, soil total P; Cmic, microbial biomass C; Nmic, microbial biomass N; Pmic, microbial biomass P; C:Nsoil, soil C to N ratio; C:Psoil, soil C to P ratio; N:Psoil, soil N to P ratio; C:Nmic, microbial biomass C to N ratio; C:Pmic, microbial biomass C to P ratio; N:Pmic, microbial biomass N to P ratio; C:Nimb, C:N stoichiometric imbalance; C:Pimb, C:P stoichiometric imbalance; N:Pimb, N:P stoichiometric imbalance; qMBC, fraction of microbial biomass C in soil C; qMBN, fraction of microbial biomass N in soil total N; qMBP, fraction of microbial biomass P in soil total P.
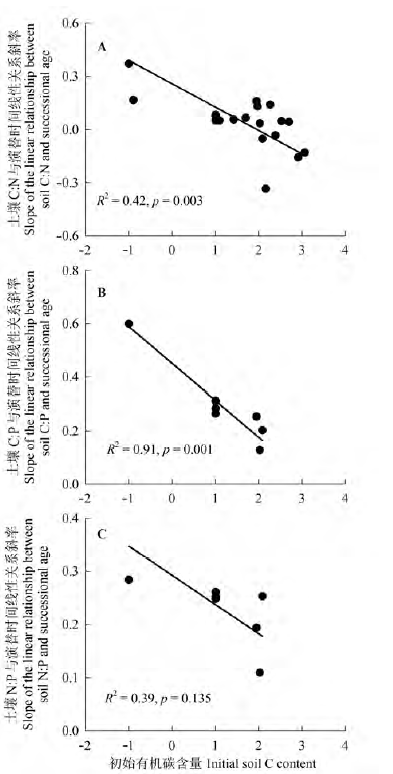
图2 演替初始有机C含量和土壤C:N:P与演替时间线性关系斜率的关系。
Fig. 2 Correlations between the initial contents of soil organic C and the slopes of linear relationships between soil C:N:P and successional age.
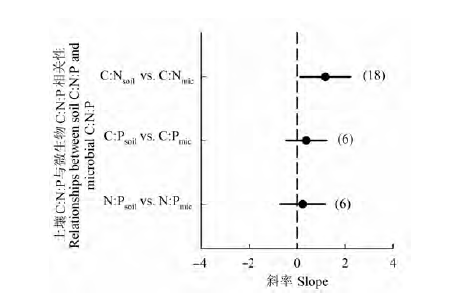
图3 不同演替序列土壤C:N:P和微生物生物量C:N:P线性关系斜率的平均值和95%置信区间。图中点和误差线分别表示斜率的平均值和95%置信区间。纵坐标括号内数字表示报道该变量的演替序列个数。C:Nsoil vs. C:Nmic, 土壤C:N与微生物C:N的线性关系; C:Psoil vs. C:Pmic, 土壤C:P与微生物C:P的线性关系; N:Psoil vs. N:Pmic, 土壤N:P与微生物N:P的线性关系。
Fig. 3 Means and 95% confidence interval of the slopes of the linear relationships between soil C:N:P and microbial C:N:P. The dots and error bars represent the means and 95% confidence intervals, respectively; the values in brackets are the numbers of age sequences. C:Nsoil vs. C:Nmic, relationship between soil C:N and microbial C:N; C:Psoil vs. C:Pmic, relationship between soil C:P and microbial C:P; N:Psoil vs. N:Pmic, relationship between soil N:P and microbial N:P.
| 分组 Group | 因子 Factor | 平方和 Sum of square | df | F | p | 方差解释率 Explanatory rate of variance (%) |
|---|---|---|---|---|---|---|
| C:N | 演替时间 Successional age | 1.8 | 1 | 28.1 | <0.001 | 2 |
| C:N化学计量不平衡性 C:N stoichiometric imbalance | 31.9 | 1 | 484.8 | <0.001 | 37 | |
| 个体序列 Individual sequence | 46.5 | 17 | 41.6 | <0.001 | 54 | |
| 误差 Error | 6.2 | 95 | 7 | |||
| C:P | 演替时间 Successional age | 0.4 | 1 | 15.1 | <0.001 | 5 |
| C:P化学计量不平衡性 C:P stoichiometric imbalance | 3.7 | 1 | 157.6 | <0.001 | 57 | |
| 个体序列 Individual sequence | 1.7 | 5 | 14.3 | <0.001 | 26 | |
| 误差 Error | 0.8 | 34 | 12 | |||
| N:P | 演替时间 Successional age | 0.4 | 1 | 9.8 | 0.004 | 5 |
| C:N化学计量不平衡性 C:N stoichiometric imbalance | 2.7 | 1 | 73.2 | <0.001 | 41 | |
| 个体序列 Individual sequence | 2.3 | 5 | 12.6 | <0.001 | 35 | |
| 误差 Error | 1.2 | 34 | 19 |
表2 广义线性模型分析演替时间、化学计量不平衡性和个体序列对微生物熵影响
Table 2 Summary of the results from a general linear model showing the integrative effects of successional ages, stoichiometric imbalances and individual sequence on microbial quotient
| 分组 Group | 因子 Factor | 平方和 Sum of square | df | F | p | 方差解释率 Explanatory rate of variance (%) |
|---|---|---|---|---|---|---|
| C:N | 演替时间 Successional age | 1.8 | 1 | 28.1 | <0.001 | 2 |
| C:N化学计量不平衡性 C:N stoichiometric imbalance | 31.9 | 1 | 484.8 | <0.001 | 37 | |
| 个体序列 Individual sequence | 46.5 | 17 | 41.6 | <0.001 | 54 | |
| 误差 Error | 6.2 | 95 | 7 | |||
| C:P | 演替时间 Successional age | 0.4 | 1 | 15.1 | <0.001 | 5 |
| C:P化学计量不平衡性 C:P stoichiometric imbalance | 3.7 | 1 | 157.6 | <0.001 | 57 | |
| 个体序列 Individual sequence | 1.7 | 5 | 14.3 | <0.001 | 26 | |
| 误差 Error | 0.8 | 34 | 12 | |||
| N:P | 演替时间 Successional age | 0.4 | 1 | 9.8 | 0.004 | 5 |
| C:N化学计量不平衡性 C:N stoichiometric imbalance | 2.7 | 1 | 73.2 | <0.001 | 41 | |
| 个体序列 Individual sequence | 2.3 | 5 | 12.6 | <0.001 | 35 | |
| 误差 Error | 1.2 | 34 | 19 |
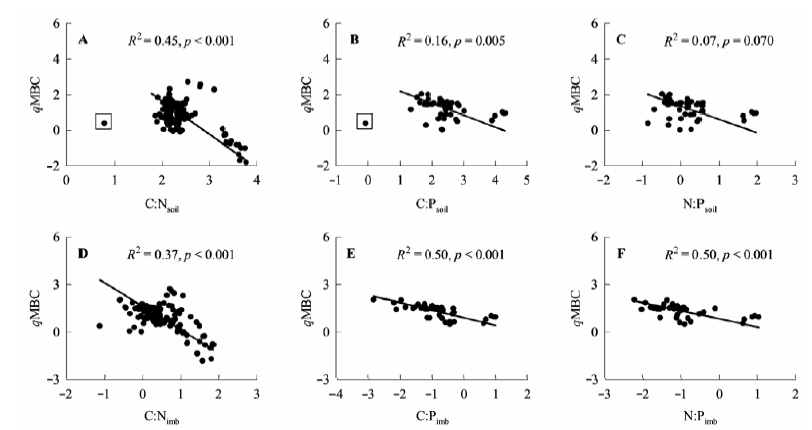
图4 微生物熵(qMBC)和土壤C:N:P以及C:N:P化学计量不平衡性的关系。C:Nsoil, 土壤C:N; C:Psoil, 土壤C:P; N:Psoil, 土壤N:P; C:Nimb, C:N化学计量不平衡性; C:Pimb, C:P化学计量不平衡性; N:Pimb, N:P化学计量不平衡性。方框中数据点为异常值。
Fig. 4 Relationships between microbial quotient (qMBC) and soil C:N:P and C:N:P stoichiometric imbalances. C:Nsoil, soil C to N ratio; C:Psoil, soil C to P ratio; N:Psoil, soil N to P ratio; C:Nimb, C:N stoichiometric imbalance; C:Pimb, C:P stoichiometric imbalance; N:Pimb, N:P stoichiometric imbalance. The dots in the black box are outliers.
| 1 | An SS, Huang YM, Zheng FL (2009). Evaluation of soil microbial indices along a revegetation chronosequence in grassland soils on the Loess Plateau, Northwest China.Applied Soil Ecology, 41, 286-292. |
| 2 | Anderson TH, Domsch K (1990). Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology & Biochemistry, 22, 251-255. |
| 3 | Banning NC, Grant CD, Jones DL, Murphy DV (2008). Recovery of soil organic matter, organic matter turnover and nitrogen cycling in a post-mining forest rehabilitation chronosequence.Soil Biology & Biochemistry, 40, 2021-2031. |
| 4 | Cao CY, Jiang DM, Teng XH, Jiang Y, Liang WJ, Cui ZB (2008). Soil chemical and microbiological properties along a chronosequence ofCaragana microphylla Lam. plantations in the Horqin sandy land of Northeast China. Applied Soil Ecology, 40, 78-85. |
| 5 | Cleveland CC, Liptzin D (2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?Biogeochemistry, 85, 235-252. |
| 6 | Elser JJ, Acharya K, Kyle M, Cotner J, Makino W, Markow T, Watts T, Hobbie S, Fagan W, Schade J, Hood J, Sterner RW (2003). Growth rate-stoichiometry couplings in diverse biota.Ecology Letters ,6, 936-943. |
| 7 | Fanin N, Fromin N, Buatois B, Hättenschwiler S (2013). An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system.Ecology Letters, 16, 764-772. |
| 8 | Fierer N, Strickland MS, Liptzin D, Bradford MA, Cleveland CC (2009). Global patterns in belowground communities.Ecology Letters, 12, 1238-1249. |
| 9 | Hu N, Li H, Tang Z, Li ZF, Li GC, Jiang Y, Hu XM, Lou YL (2016). Community size, activity and C:N stoichiometry of soil microorganisms following reforestation in a karst region.European Journal of Soil Biology, 73, 77-83. |
| 10 | Insam H, Domsch KH (1988). Relationship between soil organic carbon and microbial biomass on chronosequences of reclamation sites.Microbial Ecology, 15, 177-188. |
| 11 | Jia GM (2006). The Effects of Vegetation Succession and Land Management on Soil Nutrient, Activity and Structure of Microbial Community in Loess Plateau of Northwest China. PhD dissertation, Lanzhou University, Lanzhou.(in Chinese with English abstract)[贾国梅 (2006). 黄土高原地区植被演替和土地管理对土壤养分、微生物活性和群落结构的影响. 博士学位论文, 兰州大学, 兰州.] |
| 12 | Jia GM, Cao J, Wang CY, Wang G (2005). Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, Northwest China.Forest Ecology & Management, 217, 117-125. |
| 13 | Jiang JP, Xiong YC, Jia Y, Li FM, Xu JZ, Jiang HM (2007). Soil quality dynamics under successional alfalfa field in the semi-arid Loess Plateau of northwestern China.Arid Land Research & Management, 21, 287-303. |
| 14 | Jiang JP, Xiong YC, Jiang HM, Ye DY, Song YJ, Li FM (2009). Soil microbial activity during secondary vegetation succession in semiarid abandoned lands of Loess Plateau.Pedosphere, 19, 735-747. |
| 15 | Li P, Han WX, Yang YH, Fang JY (2014). Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems. Global Ecology & Biogeography, 23, 979-987. |
| 16 | Li Y, Wu JS, Liu SL, Shen JL, Huang DY, Su YR, Wei WW, Syers JK (2012). Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Global Biogeochemical Cycles, 26, GB4002. doi: 10.1029/2012GB004399. |
| 17 | Liu D, Huang YM, An SS (2012). Changes in soil nitrogen and microbial activity duringRobinia pseudoacacia recovery period in the Loess Hilly-Gully region. Chinese Journal of Eco-Agriculture, 20, 322-329.(in Chinese with English abstract)[刘栋, 黄懿梅, 安韶山 (2012). 黄土丘陵区人工刺槐林恢复过程中土壤氮素与微生物活性的变化. 中国生态农业学报, 20, 322-329.] |
| 18 | Liu Y, Zheng FL, An SS, He WX, Guo M, Lü CH (2010). Soil microbial biomass characteristics in response to vegetation restoration on abandoned lands in Yangou Watershed of China.Plant Nutrition & Fertilizer Science, 16, 824-832.(in Chinese with English abstract)[刘雨, 郑粉莉, 安韶山, 和文祥, 郭曼, 吕春华 (2010). 燕沟流域土壤微生物学性质对植被恢复过程的响应. 植物营养与肥料学报, 16, 824-832.] |
| 19 | Liu YM, Li XR, Xing ZS, Zhao X, Pan YX (2013). Responses of soil microbial biomass and community composition to biological soil crusts in the revegetated areas of the Tengger Desert.Applied Soil Ecology, 65, 52-59. |
| 20 | McGroddy ME, Daufresne T, Hedin LO (2004). Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios.Ecology, 85, 2390-2401. |
| 21 | Mooshammer M, Wanek W, Zechmeister-Boltenstern S, Richter A (2014). Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources.Frontiers in Microbiology, 5, 22. doi: 10.3389/ fmicb.2014.00022. |
| 22 | Odum EP (1969). The strategy of ecosystem development.Science, 164, 262-270. |
| 23 | Redfield AC (1958). The biological control of chemical factors in the environment.American Scientist, 64, 205-221. |
| 24 | Shi XM, Qi JH, Song L, Liu WY, Huang JB, Li S, Lu HZ, Chen X (2015). C, N and P stoichiometry of two dominant seedlings and their responses to nitrogen additions in the montane moist evergreen broad-leaved forest in Ailao Mountains, Yunnan.Chinese Journal of Plant Ecology, 39, 962-970.(in Chinese with English abstract)[石贤萌, 杞金华, 宋亮, 刘文耀, 黄俊彪, 李苏, 卢华正, 陈曦 (2015). 哀牢山中山湿性常绿阔叶林两种优势幼苗C、N、P化学计量特征及其对N沉降增加的响应. 植物生态学报, 39, 962-970.] |
| 25 | Singh KP, Mandal TN, Tripathi SK (2001). Patterns of restoration of soil physciochemical properties and microbial biomass in different landslide sites in the sal forest ecosystem of Nepal Himalaya.Ecological Engineering, 17, 385-401. |
| 26 | Spohn M, Novak TJ, Incze J, Giani L (2016). Dynamics of soil carbon, nitrogen, and phosphorus in calcareous soils after land-use abandonment—A chronosequence study.Plant & Soil, 401, 185-196. |
| 27 | Sterner RW, Elser JJ (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton, USA. |
| 28 | Tian HQ, Chen GS, Zhang C, Melillo JM, Hall CAS (2010). Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data.Biogeochemistry, 98, 139-151. |
| 29 | Vergutz L, Manzoni S, Porporato A, Novais RF, Jackson RB (2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants.Ecological Monographs, 82, 205-220. |
| 30 | Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010). Terrestrial phosphorus limitation: Mechanisms implica- tions and nitrogen-phosphorus interactions.Ecological Applications, 20, 5-15. |
| 31 | Warton DI, Wright IJ, Falster DS, Westoby M (2006). Bivariate line-fitting methods for allometry.Biological Reviews, 81, 259-291. |
| 32 | Weedon JT, Cornwell WK, Cornelissen JHC, Zanne AE, Wirth C, Coomes DA (2009). Global meta-analysis of wood decomposition rates: A role for trait variation among tree species?Ecology Letters, 12, 45-56. |
| 33 | Xiao L, Liu GB, Xue S, Zhang C (2013). Soil microbial community composition during natural recovery in the Loess Plateau, China.Journal of Integrative Agriculture, 12, 1872-1883. |
| 34 | Xu XF, Schimel JP, Thornton PE, Song X, Yuan FM, Goswami S (2014). Substrate and environmental controls on microbial assimilation of soil organic carbon: A framework for Earth system models.Ecology Letters, 17, 547-555. |
| 35 | Xu XF, Thornton PE, Post WM (2013). A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecology & Biogeography, 22, 737-749. |
| 36 | Xue S, Liu GB, Dai QH, Li XL, Wu RJ (2008). Dynamic changes of soil microbial biomass in the restoration process of shrub plantations in Loess Hilly area.Chinese Journal of Applied Ecology, 19, 517-523.(in Chinese with English abstract)[薛萐, 刘国彬, 戴全厚, 李小利, 吴瑞俊 (2008). 黄土丘陵区人工灌木林恢复过程中的土壤微生物生物量演变. 应用生态学报, 19, 517-523.] |
| 37 | Xue S, Liu GB, Dai QH, Wei W, Hou XL (2007). Evolution of soil microbial biomass in the restoration process of artificialRobinia pseudoacacia under erosion environment. Acta Ecologica Sinica, 27, 909-917.(in Chinese with English abstract)[薛萐, 刘国彬, 戴全厚, 卫伟, 侯喜禄 (2007). 侵蚀环境生态恢复过程中人工刺槐林(Robinia pseudoacacia)土壤微生物量演变特征. 生态学报, 27, 909-917.] |
| 38 | Xue S, Liu GB, Dai QH, Zhang C, Yu N (2009). Dynamics of soil microbial biomass on the abandoned cropland in Loess Hilly area.Scientia Agricultura Sinica, 42, 943-950.(in Chinese with English abstract)[薛萐, 刘国彬, 戴全厚, 张超, 余娜 (2009). 黄土丘陵区退耕撂荒地土壤微生物量演变过程. 中国农业科学, 42, 943-950.] |
| 39 | Yang N, Zou DS, Yang MY, Lin ZG, Song GT, Chen ZY, Zhao LF (2014). Changes of soil properties in re-vegetation stages on sloping-land with purple soils in Hengyang of Hunan Province, South-central China.Acta Ecologica Sinica, 34, 2693-2701.(in Chinese with English abstract)[杨宁, 邹冬生, 杨满元, 林仲桂, 宋光桃, 陈志阳, 赵林峰 (2014). 衡阳紫色土丘陵坡地植被恢复阶段土壤特性的演变. 生态学报, 34, 2693-2701.] |
| 40 | Yang YH, Luo YQ (2011). Carbon : nitrogen stoichiometry in forest ecosystems during stand development.Global Ecology & Biogeography, 20, 354-361. |
| 41 | Yang YH, Luo YQ, Finzi AC (2011). Carbon and nitrogen dynamics during forest stand development: A global synthesis.New Phytologist ,190, 977-989. |
| 42 | Yuan ZY, Chen HY, Reich PB (2011). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus.Nature Communications, 2, 344. |
| 43 | Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W (2015). The application of ecological stoichiometry to plant-microbial- soil organic matter transformations.Ecological Mono- graphs, 85, 133-155. |
| 44 | Zeng DH, Chen GS (2005). Ecological stoichiometry: A science to explore the complexity of living systems.Acta Phytoecologica Sinica, 29, 1007-1019.(in Chinese with English abstract)[曾德慧, 陈广生 (2005). 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 29, 1007-1019.] |
| 45 | Zeng DP, Jiang LL, Zeng CS, Wang WQ, Wang C (2013). Reviews on the ecological stoichiometry characteristics and its applications.Acta Ecologica Sinica, 33, 5484-5492.(in Chinese with English abstract)[曾冬萍, 蒋利玲, 曾从盛, 王维奇, 王纯 (2013). 生态化学计量学特征及其应用研究进展. 生态学报, 33, 5484-5492.] |
| 46 | Zhou ZH, Wang CK (2015). Reviews and syntheses: Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China’s forest ecosystems.Biogeosciences, 12, 6751-6760. |
| 47 | Zhou ZH, Wang CK (2016). Responses and regulation mechanisms of microbial decomposers to substrate carbon, nitrogen, and phosphorus stoichiometry.Chinese Journal of Plant Ecology ,40, 620-630.(in Chinese with English abstract)[周正虎, 王传宽 (2016). 微生物对分解底物碳氮磷化学计量的响应和调节机制. 植物生态学报, 40, 620-630.] |
| 48 | Zhou ZH, Wang CK (2017). Soil-microbe-mineralization car- bon and nitrogen stoichiometry under different land-uses in the Maoershan region. Acta Ecologica Sinica, 37, in press. doi: 10.5846/stxb201512242569.(in Chinese with English abstract)[周正虎, 王传宽 (2017). 帽儿山地区不同土地利用方式下土壤-微生物-矿化碳氮化学计量特征. 生态学报, 37, 待发表. doi: 10.5846/stxb201512242569.] |
| 49 | Zhou ZH, Wang CK, Zhang QZ (2015). The effect of land use change on soil carbon, nitrogen, and phosphorus contents and their stoichiometry in temperate sapling stands in northeastern China.Acta Ecologica Sinica, 35, 6694-6702.(in Chinese with English abstract)[周正虎, 王传宽, 张全智 (2015). 土地利用变化对东北温带幼龄林土壤碳氮磷含量及其化学计量特征的影响. 生态学报, 35, 6694-6702.] |
| 50 | Zhu HH, He XY, Wang KL, Su YR, Wu JS (2012). Interactions of vegetation succession, soil bio-chemical properties and microbial communities in a Karst ecosystem.European Journal of Soil Biology, 51, 1-7. |
| [1] | 俞庆水 倪晓凤 吉成均 朱江玲 唐志尧 方精云. 10年氮磷添加对海南尖峰岭两种热带雨林优势植物叶片非结构性碳水化合物的影响[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 彭仲韬 金光泽 刘志理. 小兴安岭三种槭树叶性状随植株大小和林冠条件的变异[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [3] | 刘瑶 钟全林 徐朝斌 程栋梁 郑跃芳 邹宇星 张雪 郑新杰 周云若. 不同大小刨花楠细根功能性状与根际微环境关系[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [4] | 周建 王焓. 森林径级结构研究:从统计描述到理论演绎[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [5] | 张智洋 赵颖慧 甄贞. 1986-2022年松花江流域陆地生态系统碳储量动态监测[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [6] | 张文瑾 佘维维 秦树高 乔艳桂 张宇清. 氮和水分添加对黑沙蒿群落优势植物叶片氮磷化学计量特征的影响[J]. 植物生态学报, 2024, 48(5): 590-600. |
| [7] | 王袼, 胡姝娅, 李阳, 陈晓鹏, 李红玉, 董宽虎, 何念鹏, 王常慧. 不同类型草原土壤净氮矿化速率的温度敏感性[J]. 植物生态学报, 2024, 48(4): 523-533. |
| [8] | 张计深, 史新杰, 刘宇诺, 吴阳, 彭守璋. 气候变化下中国潜在自然植被生态系统碳储量动态[J]. 植物生态学报, 2024, 48(4): 428-444. |
| [9] | 秦文宽, 张秋芳, 敖古凯麟, 朱彪. 土壤有机碳动态对增温的响应及机制研究进展[J]. 植物生态学报, 2024, 48(4): 403-415. |
| [10] | 黄玲, 王榛, 马泽, 杨发林, 李岚, SEREKPAYEV Nurlan, NOGAYEV Adilbek, 侯扶江. 长期放牧和氮添加对黄土高原典型草原长芒草种群生长的影响[J]. 植物生态学报, 2024, 48(3): 317-330. |
| [11] | 杨安娜, 李曾燕, 牟凌, 杨柏钰, 赛碧乐, 张立, 张增可, 王万胜, 杜运才, 由文辉, 阎恩荣. 上海大金山岛不同植被类型土壤细菌群落的变异[J]. 植物生态学报, 2024, 48(3): 377-389. |
| [12] | 牛一迪, 蔡体久. 大兴安岭北部次生林演替过程中物种多样性的变化及其影响因子[J]. 植物生态学报, 2024, 48(3): 349-363. |
| [13] | 吴君梅, 曾泉鑫, 梅孔灿, 林惠瑛, 谢欢, 刘苑苑, 徐建国, 陈岳民. 土壤磷有效性调控亚热带森林土壤酶活性和酶化学计量对凋落叶输入的响应[J]. 植物生态学报, 2024, 48(2): 242-253. |
| [14] | 韩大勇, 李海燕, 张维, 杨允菲. 松嫩草地全叶马兰种群分株养分的季节运转及衰老过程[J]. 植物生态学报, 2024, 48(2): 192-200. |
| [15] | 颜辰亦, 龚吉蕊, 张斯琦, 张魏圆, 董学德, 胡宇霞, 杨贵森. 氮添加对内蒙古温带草原土壤活性有机碳的影响[J]. 植物生态学报, 2024, 48(2): 229-241. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19