
植物生态学报 ›› 2017, Vol. 41 ›› Issue (9): 985-994.DOI: 10.17521/cjpe.2017.0005
李群, 赵成章*( ), 赵连春, 王建良, 张伟涛, 姚文秀
), 赵连春, 王建良, 张伟涛, 姚文秀
收稿日期:2017-01-07
修回日期:2017-07-09
出版日期:2017-09-10
发布日期:2017-10-23
通讯作者:
赵成章
基金资助:
Qun LI, Cheng-Zhang ZHAO*(), Lian-Chun ZHAO, Jian-Liang WANG, Wei-Tao ZHANG, Wen-Xiu YAO
Received:2017-01-07
Revised:2017-07-09
Online:2017-09-10
Published:2017-10-23
Contact:
Cheng-Zhang ZHAO
摘要:
植物比叶面积(SLA)与叶片热耗散的关联性分析, 对于探究植物光合产物积累与耗散及其适应特殊生境有重要意义。该研究以秦王川国家湿地公园芦苇(Phragmites australis)种群为研究对象, 根据距离水域的远近分别依次设置3个试验样地: I (离水域25-40 m, 土壤含水量(SM) 28.94%)、II (离水域10-25 m, SM 45.97%)、III (离水域0-10 m, SM 76.81%), 研究了秦王川盐沼湿地芦苇SLA与叶片热耗散的关系。结果表明: 随着土壤含水量的增加, 湿地植被群落的高度和地上生物量呈现逐渐增加的趋势, 土壤电导率和光合有效辐射呈显著减小的趋势; 叶面积、净光合速率和蒸腾速率均呈增加的趋势, 叶厚度呈减少的趋势, SLA呈现先增加后减小的趋势, 叶干质量呈现先减少后增加的趋势; 从样地I到样地III, 调节性能量耗散的量子产额(Y(NPQ))、非光化学淬灭系数呈减少的趋势, 实际光合效率、光化学淬灭系数呈增加的趋势, 非调节性能量耗散的量子产额呈先增加后减少的趋势; 在样地I和III, 芦苇SLA与Y(NPQ)呈极显著负相关关系(p < 0.01), 在样地II, SLA与Y(NPQ)显著负相关(p < 0.05)。随土壤含水量的梯度性变化, 芦苇种群通过调整叶片构建模式从而适时调整SLA的大小以改变叶片热耗散, 实现对植物吸收光能的有效利用, 反映了植物为适应特殊生境通过改变表型调控叶片热耗散实现自我保护的机制。
李群, 赵成章, 赵连春, 王建良, 张伟涛, 姚文秀. 秦王川盐沼湿地芦苇比叶面积与叶片热耗散的关联性分析. 植物生态学报, 2017, 41(9): 985-994. DOI: 10.17521/cjpe.2017.0005
Qun LI, Cheng-Zhang ZHAO, Lian-Chun ZHAO, Jian-Liang WANG, Wei-Tao ZHANG, Wen-Xiu YAO. Empirical relationship between specific leaf area and thermal dissipation of Phragmites australis in salt marshes of Qinwangchuan. Chinese Journal of Plant Ecology, 2017, 41(9): 985-994. DOI: 10.17521/cjpe.2017.0005
| 样地 Plot | 土壤含水量 Soil moisture content (%) | 土壤电导率 Soil electrical conductivity (ms·cm-1) | PAR (μmol·m-2·s-1) | 高度 Height (cm) | 地上生物量 Aboveground biomass (g·m-2) |
|---|---|---|---|---|---|
| I | 28.94 ± 1.45c | 2.44 ± 0.12a | 1 236.3 ± 3.78c | 141.80 ± 7.09c | 1 088.12 ± 54.40c |
| II | 45.97 ± 2.30b | 1.85 ± 0.09b | 866.0 ± 4.43b | 191.60 ± 9.58b | 1 759.36 ± 87.97b |
| III | 76.81 ± 3.84a | 0.65 ± 0.03c | 587.0 ± 4.99a | 328.80 ± 16.64a | 3 195.32 ± 159.77a |
表1 不同样地的湿地群落生物学特征和土壤特性(平均值±标准误差)
Table 1 Physiological and soil characteristics of wetland community in three sampling plots (mean ± SE)
| 样地 Plot | 土壤含水量 Soil moisture content (%) | 土壤电导率 Soil electrical conductivity (ms·cm-1) | PAR (μmol·m-2·s-1) | 高度 Height (cm) | 地上生物量 Aboveground biomass (g·m-2) |
|---|---|---|---|---|---|
| I | 28.94 ± 1.45c | 2.44 ± 0.12a | 1 236.3 ± 3.78c | 141.80 ± 7.09c | 1 088.12 ± 54.40c |
| II | 45.97 ± 2.30b | 1.85 ± 0.09b | 866.0 ± 4.43b | 191.60 ± 9.58b | 1 759.36 ± 87.97b |
| III | 76.81 ± 3.84a | 0.65 ± 0.03c | 587.0 ± 4.99a | 328.80 ± 16.64a | 3 195.32 ± 159.77a |
| 样地 Plot | I | II | III |
|---|---|---|---|
| 叶面积 Leaf area (cm2) | 7.71 ± 0.36c | 14.19 ± 0.71b | 28.75 ± 1.44a |
| 叶厚度 Leaf thickness (mm) | 0.36 ± 0.02a | 0.32 ± 0.02b | 0.27 ± 0.01c |
| 叶干质量 Leaf dry mass (g) | 0.39 ± 0.02b | 0.22 ± 0.01c | 0.72 ± 0.04a |
| 比叶面积 Specific leaf area (cm2·g-1) | 19.77 ± 0.99c | 64.50 ± 3.23a | 39.93 ± 2.00b |
| Pn (μmol CO2·m-2 ·s-1) | 3.39 ± 0.17c | 4.41 ± 0.22b | 7.34 ± 0.37a |
| Tr (mmol H2O·m-2·s-1) | 1.00 ± 0.05b | 1.04 ± 0.05b | 1.17 ± 0.06a |
表2 不同样地芦苇叶性状与光合生理参数特征(平均值±标准误差)
Table 2 Leaf characteristics and physiological measures of Phragmites australis in different plots (mean ± SE)
| 样地 Plot | I | II | III |
|---|---|---|---|
| 叶面积 Leaf area (cm2) | 7.71 ± 0.36c | 14.19 ± 0.71b | 28.75 ± 1.44a |
| 叶厚度 Leaf thickness (mm) | 0.36 ± 0.02a | 0.32 ± 0.02b | 0.27 ± 0.01c |
| 叶干质量 Leaf dry mass (g) | 0.39 ± 0.02b | 0.22 ± 0.01c | 0.72 ± 0.04a |
| 比叶面积 Specific leaf area (cm2·g-1) | 19.77 ± 0.99c | 64.50 ± 3.23a | 39.93 ± 2.00b |
| Pn (μmol CO2·m-2 ·s-1) | 3.39 ± 0.17c | 4.41 ± 0.22b | 7.34 ± 0.37a |
| Tr (mmol H2O·m-2·s-1) | 1.00 ± 0.05b | 1.04 ± 0.05b | 1.17 ± 0.06a |
| 样地 Plot | Y(II) | QP | NPQ | Y(NO) | Y(NPQ) |
|---|---|---|---|---|---|
| I | 0.21 ± 0.01c | 0.42 ± 0.02b | 0.45 ± 0.02a | 0.29 ± 0.01a | 0.50 ± 0.03a |
| II | 0.25 ± 0.01b | 0.60 ± 0.03a | 0.43 ± 0.02a | 0.30 ± 0.02a | 0.45 ± 0.02b |
| III | 0.36 ± 0.02a | 0.65 ± 0.03a | 0.34 ± 0.02b | 0.28 ± 0.01a | 0.36 ± 0.02c |
表3 不同样地芦苇叶片叶绿素荧光特征(平均值±标准误差)
Table 3 Leaf chlorophyll fluorescences of Phragmites australis in different plots (mean ± SE)
| 样地 Plot | Y(II) | QP | NPQ | Y(NO) | Y(NPQ) |
|---|---|---|---|---|---|
| I | 0.21 ± 0.01c | 0.42 ± 0.02b | 0.45 ± 0.02a | 0.29 ± 0.01a | 0.50 ± 0.03a |
| II | 0.25 ± 0.01b | 0.60 ± 0.03a | 0.43 ± 0.02a | 0.30 ± 0.02a | 0.45 ± 0.02b |
| III | 0.36 ± 0.02a | 0.65 ± 0.03a | 0.34 ± 0.02b | 0.28 ± 0.01a | 0.36 ± 0.02c |
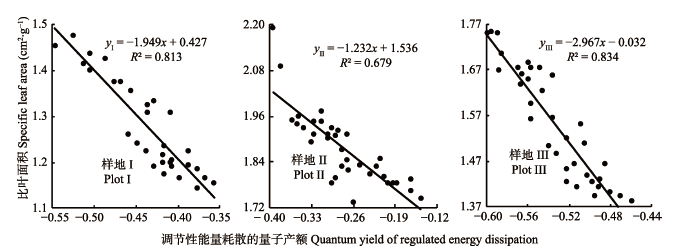
图1 不同样地芦苇比叶面积(SLA)与调节性能量耗散的量子产额(Y(NPQ))之间关系。
Fig. 1 Relationship between specific leaf area (SLA) and quantum yield of regulated energy dissipation (Y(NPQ)) of Phragmites australis at three sampling plots.
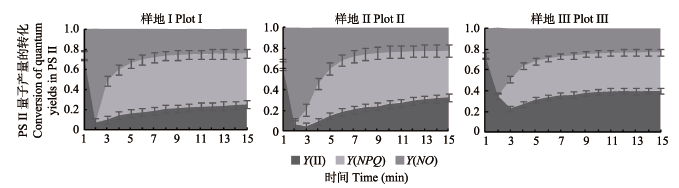
图2 不同样地芦苇叶片中PSII量子产量的转化(平均值±标准偏差)。Y(II), 光系统II光化学量子产量; Y(NO), PSII中荧光和不依赖光的基础热耗散量子产量; Y(NPQ), PSII中用于热耗散的调节性能量耗散的量子产量。光合有效辐射= 1β200 μmol· m-2·s-1 。
Fig. 2 Changes in quantum yields (PSII) of reed leaves at three sampling plots (mean ± SD). Y(II), photochemical quantum yields in PSII; Y(NO), quantum yield of fluorescence and light-independent constitution thermal dissipation; Y(NPQ), quantum yield of thermal dissipation used in regulatory energy dissipation. Photosynthetically active radiation = 1β200 μmol·m-2·s-1.
| [1] | Amoros-Lopez J, Gomez-Chova L, Vila-Frances J, Alonso L, Calpe J, Moreno J, del Valle-Tascon S (2008). Evaluation of remote sensing of vegetation fluorescence by the analysis of diurnal cycles.International Journal of Remote Sensing, 29, 5423-5436. |
| [2] | Baker NR (1991). A possible role for photosystem II in environmental perturbations of photosynthesis.Physiologia Plantarum, 81, 563-570. |
| [3] | Dang JJ, Zhao CZ, Li J, Hou ZJ, Dong XG (2015). Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China. Chinese Journal of Plant Ecology, 39, 23-31. (in Chinese with English abstract)[党晶晶, 赵成章, 李钰, 侯兆疆, 董小刚 (2015). 祁连山高寒草地甘肃臭草叶性状与坡向间的关系. 植物生态学报, 39, 23-31.] |
| [4] | Demmig-Adams B, Adams WW III (2006). Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation.New Phytologist, 172, 11-21. |
| [5] | Deng JM, Li T, Wang GX, Liu J, Yu ZL, Zhao CM, Ji MF, Zhang Q (2008). Trade-offs between the metabolic rate and population density of plants.PLOS ONE, 3, e1799. doi: 10.1371/journal.pone.0001799. |
| [6] | Givnish TJ (1978). Ecological aspects of plant morphology: Leaf form in relation to environment.Acta Biotheoretica, 27, 83-142. |
| [7] | Gong JR, Huang YM, Ge ZW, Duan QW, You X, An R, Zhang XS (2009). Ecological responses to soil water content in four hybridPopulous clones. Chinese Journal of Plant Ecology, 33, 387-396. (in Chinese with English abstract)[龚吉蕊, 黄永梅, 葛之葳, 段庆伟, 尤鑫, 安然, 张新时 (2009). 4种杂交杨对土壤水分变化的生态学响应. 植物生态学报, 33, 387-396.] |
| [8] | Gong R, Gao Q (2015). Research progress in the effects of leaf hydraulic characteristics on plant physiological functions.Chinese Journal of Plant Ecology, 39, 300-308. (in Chinese with English abstract)[龚容, 高琼 (2015). 叶片结构的水力学特性对植物生理功能影响的研究进展. 植物生态学报, 39, 300-308.] |
| [9] | Han L, Zhao CZ, Xu T, Feng W, Duan BB, Zheng HL (2016). Trade-off between leaf size and vein density of Achnatherum splendens in Zhangye wetland. Chinese Journal of Plant Ecology, 40, 788-797. (in Chinese with English abstract)[韩玲, 赵成章, 徐婷, 冯威, 段贝贝, 郑慧玲 (2016). 张掖湿地芨芨草叶大小和叶脉密度的权衡关系. 植物生态学报, 40, 788-797.] |
| [10] | Hu WH, Zhang SS, Xiao YA, Yan XH (2015). Physiological responses and photo-protective mechanisms of two Rhododendron plants to natural sunlight after long term shading. Chinese Journal of Plant Ecology, 39, 1093-1100. (in Chinese with English abstract)[胡文海, 张斯斯, 肖宜安, 闫小红 (2015). 两种杜鹃花属植物对长期遮阴后全光照环境的生理响应及其光保护机制. 植物生态学报, 39, 1093-1100.] |
| [11] | Hu YS, Mo XY, Liu YH (2015). Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains.Acta Ecologica Sinica, 35, 1480-1487. (in Chinese with English abstract)[胡耀升, 么旭阳, 刘艳红 (2015). 长白山森林不同演替阶段比叶面积及其影响因子. 生态学报, 35, 1480-1487.] |
| [12] | Hultine KR, Marshall JD (2000). Altitude trends in conifer leaf morphology and stable carbon isotope composition.Oecologia, 123, 32-40. |
| [13] | Kang SZ, Zhang JH (2004). Controlled alternate partial root- zone irrigation: Its physiological consequences and impact on water use efficiency.Journal of Experimental Botany, 55, 2437-2446. |
| [14] | Knight CA, Ackerly DD (2003). Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size: Congeneric species from desert and coastal environments. New Phytologist, 160, 337-347. |
| [15] | Komura M, Yamagishi A, Shibata Y, Iwasaki I, Itoh S (2010). Mechanism of strong quenching of photosystem II chlorophyll fluorescence under drought stress in a lichen,Physciella melanchra, studied by subpicosecond fluorescence spectroscopy. Biochim Biophys Acta, 1797, 331-338. |
| [16] | Kramer DM, Johnson G, Kiirats O, Edwards GE (2004). New fluorescence parameters for the determination of QA redox state and excitation energy fluxes.Photosynthesis Research, 79, 209-218. |
| [17] | Li L, Zhou DW, Sheng LX (2011). Density dependence-determined plant biomass allocation pattern. Chinese Journal of Ecology, 30, 1579-1589. (in Chinese with English abstract)[黎磊, 周道玮, 盛连喜 (2011). 密度制约决定的植物生物量分配格局. 生态学杂志, 30, 1579-1589.] |
| [18] | Li R, Jiang ZM, Zhang SX, Cai J (2015). A review of new research progress on the vulnerability of xylem embolism of woody plants. Chinese Journal of Plant Ecology, 39, 838-848. (in Chinese with English abstract)[李荣, 姜在民, 张硕新, 蔡靖 (2015). 木本植物木质部栓塞脆弱性研究新进展. 植物生态学报, 39, 838-848.] |
| [19] | Li YL, Cui JY, Su YZ (2005). Specific leaf area and leaf dry matter content of some plants in different dune habitats.Acta Ecologica Sinica, 25, 304-311. (in Chinese with English abstract)[李玉霖, 崔建垣, 苏永中 (2005). 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 25, 304-311.] |
| [20] | Liu MH, Xin ZM, Xu J, Sun F, Dou LJ, li YH (2013). Influence of leaf size of plant on leaf transpiration and temperature in arid regions.Chinese Journal of Plant Ecology, 37, 436-442. (in Chinese with English abstract)[刘明虎, 辛智鸣, 徐军, 孙非, 窦立军, 李永华 (2013). 干旱区植物叶片大小对叶表面蒸腾及叶温的影响. 植物生态学报, 37, 436-442.] |
| [21] | Luo XG (2003). The discussion and analysis of Qinwangchuan irrigated area drainage and salinization prevention question.The Agriculture of Gansu, (12), 89. (in Chinese)[罗星光 (2003). 秦王川灌区排水和盐渍化防治问题的探讨与分析. 甘肃农业, (12), 89.] |
| [22] | Ogburn RM, Edwards EJ (2012). Quantifying succulence: A rapid, physiologically meaningful metric of plant water storage.Plant, Cell & Environment, 35, 1533-1542. |
| [23] | Poorter L, Bongers F (2006). Leaf traits are good predictors of plant performance across 53 rain forest species.Ecology, 87, 1733-1743. |
| [24] | Qi ZW, Jiang N, Gao YN, Li Y, Wang KY, Zhang C (2016). Responses of photosynthesis of Phragmites australis to water-salt indicators of soil in Dongtan wetlands, Chongming Island. Wetland Island, 14, 538-545. (in Chinese with English abstract)[戚志伟, 姜楠, 高艳娜, 李艳, 王开运, 张超 (2016). 崇明岛东滩湿地芦苇光合作用对土壤水盐因子的响应. 湿地科学, 14, 538-545.] |
| [25] | Ren QJ, Li HL, Pu HY (2015). Comparison of physiological and leaf morphological traits for photosynthesis of the 51 plant species in the Maqu alpine swamp meadow.Chinese Journal of Plant Ecology, 39, 593-603. (in Chinese with English abstract)[任青吉, 李宏林, 卜海燕 (2015). 玛曲高寒沼泽化草甸51种植物光合生理和叶片形态特征的比较. 植物生态学报, 39, 593-603.] |
| [26] | Rozema J (1996). Biology of Halophytes: Halophytes and Biosaline Agriculture. Marcel Dekker, New York. 17-30. |
| [27] | Schreiber U (2004). Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: An overview. In: Papageorgiou GC, Govindjee eds. Chlorophyll a Fluorescence. Springer, Heidelberg, Germany. 279-319. |
| [28] | Schreiber U, Bilger W, Neubauer C (1994). Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In: Schulze ED, Caldwell MM eds. Ecophysiology of Photosynthesis. Springer-Verlag, Berlin. 49-70. |
| [29] | Scoffoni C, Rawls M, McKown A, Cochard H, Sack L (2011). Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture.Plant Physiology, 156, 832-843. |
| [30] | Shi Y, Wen ZM, Gong SH, Song G, Zhen Y, Ding M (2012). Trait variations along a climatic gradient in hilly area of Loess Plateau.Research of Soil and Water Conservation, 19(1), 107-111, 116. (in Chinese with English abstract)[施宇, 温仲明, 龚时慧, 宋光, 郑颖, 丁曼 (2012). 黄土丘陵区植物功能性状沿气候梯度的变化规律. 水土保持研究, 19(1), 107-111, 116.] |
| [31] | Song LL, Fan JW, Wu SH, Zhong HP, Wang N (2012). Response characteristics of leaf traits of common species along an altitudinal gradient in Hongchiba grassland, Chongqing.Acta Ecologica Sinica, 32, 2759-2767. (in Chinese with English abstract)[宋璐璐, 樊江文, 吴绍洪, 钟华平, 王宁 (2012). 红池坝草地常见物种叶片性状沿海拔梯度的响应特征. 生态学报, 32, 2759-2767.] |
| [32] | Westoby M, Wright IJ (2003). The leaf size-twig size spectrum and its relationship to other important spectra of variation among species.Oecologia, 135, 621-628. |
| [33] | Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JH, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas ML, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004). The worldwide leaf economics spectrum.Nature, 428, 821-827. |
| [34] | Wright IJ, Westoby M, Reich PB (2002). Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span.Journal of Ecology, 90, 534-543. |
| [35] | Xiang F, Zhou Q, Tian XR, Chen GX, Xiao Y (2014). Leaf morphology and PSII chlorophyll fluorescence parameters in leaves ofSinosenecio jishouensis in different habitats. Acta Ecologica Sinica, 34, 337-344. (in Chinese with English abstract)[向芬, 周强, 田向荣, 陈功锡, 肖艳 (2014). 不同生境吉首蒲儿根叶片形态和叶绿素荧光特征的比较. 生态学报, 34, 337-344.] |
| [36] | Yu HY, Chen YT, Xu ZZ, Zhou GS (2014). Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol. Chinese Journal of Plant Ecology, 38, 1029-1040. (in Chinese with English abstract)[于鸿莹, 陈莹婷, 许振柱, 周广胜 (2014). 内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析. 植物生态学报, 38, 1029-1040.] |
| [37] | Yu WY, Ji RP, Feng Rui, Wu JW, Zhao XL, Zhang YS (2014). Advances in simulation of reed wetland evapotranspiration based on multiple time and space scales.Chinese Journal of Ecology, 33, 1388-1394. (in Chinese with English abstract)[于文颖, 纪瑞鹏, 冯锐, 武晋雯, 赵先丽, 张玉书 (2014). 芦苇湿地多时空尺度蒸散模拟研究进展. 生态学杂志, 33, 1388-1394.] |
| [38] | Zhang C, Zhan DX, Zhang PP, Zhang YL, Luo HH, Zhang WF (2014). Responses of photorespiration and thermal dissipation in PSII to soil water in cotton bracts.Chinese Journal of Plant Ecology, 38, 387-395. (in Chinese with English abstract)[张超, 占东霞, 张鹏鹏, 张亚黎, 罗宏海, 张旺锋 (2014). 棉花苞叶光呼吸和PSII热耗散对土壤水分的响应, 植物生态学报, 38, 387-395.] |
| [39] | Zhang YQ, Liang CZ, Wang W, Wang LX, Peng JT, Yan JC, Jia JZ (2010). Soil salinity andAchnatherum splendens distribution. Chinese Journal of Ecology, 29, 2438-2443. (in Chinese with English abstract)[张雅琼, 梁存柱, 王炜, 王立新, 彭江涛, 闫建成, 贾成朕 (2010). 芨芨草群落土壤盐分特征. 生态学杂志, 29, 2438-2443.] |
| [40] | Zhu QH, Xia HX, Xie HH, Li Q, Ding WQ (2013). Response ofAcorus tatarinowii in chlorophyll fluorescence to flooding stress. Oceanologia et Limnologia Sinica, 44, 1494-1499. (in Chinese with English abstract)[朱启红, 夏红霞, 谢海航, 李强, 丁武泉 (2013). 石菖蒲叶绿素荧光特性对淹水胁迫的响应. 海洋与湖沼, 44, 1494-1499.] |
| [41] | Zhuang Y, Sun YX, Wang ZS, Yang LL, Deng ZF, Yao ZG, An SQ (2010). Research advances in ecotypes ofPhragmites australis. Acta Ecologica Sinica, 30, 2173-2181. (in Chinese with English abstract)[庄瑶, 孙一香, 王中生, 杨琳璐, 邓自发, 姚志刚, 安树青 (2010). 芦苇生态型研究进展. 生态学报, 30, 2173-2181.] |
| [1] | 李雪, 董杰, 韩广轩, 张奇奇, 谢宝华, 李培广, 赵明亮, 陈克龙, 宋维民. 黄河三角洲典型滨海盐沼湿地土壤CO2和CH4排放对水盐变化的响应[J]. 植物生态学报, 2023, 47(3): 434-446. |
| [2] | 史欢欢, 雪穷, 于振林, 汪承焕. 密度、物种比例对盐沼植物种子萌发阶段种内、种间相互作用的影响[J]. 植物生态学报, 2023, 47(1): 77-87. |
| [3] | 王广亚, 陈柄华, 黄雨晨, 金光泽, 刘志理. 着生位置对水曲柳小叶性状变异及性状间相关性的影响[J]. 植物生态学报, 2022, 46(6): 712-721. |
| [4] | 韩广轩, 王法明, 马俊, 肖雷雷, 初小静, 赵明亮. 滨海盐沼湿地蓝色碳汇功能、形成机制及其增汇潜力[J]. 植物生态学报, 2022, 46(4): 373-382. |
| [5] | 靳川, 李鑫豪, 蒋燕, 徐铭泽, 田赟, 刘鹏, 贾昕, 查天山. 黑沙蒿光合能量分配组分在生长季的相对变化与调控机制[J]. 植物生态学报, 2021, 45(8): 870-879. |
| [6] | 韩广轩, 李隽永, 屈文笛. 氮输入对滨海盐沼湿地碳循环关键过程的影响及机制[J]. 植物生态学报, 2021, 45(4): 321-333. |
| [7] | 张自琰, 金光泽, 刘志理. 不同区域针叶年龄对红松叶性状及相关关系的影响[J]. 植物生态学报, 2021, 45(3): 253-264. |
| [8] | 李群, 赵成章, 王继伟, 文军, 李子琴, 马俊逸. 甘肃小苏干湖盐沼湿地盐地风毛菊叶形态-光合生理特征对淹水的响应[J]. 植物生态学报, 2019, 43(8): 685-696. |
| [9] | 郭瑞, 周际, 刘琪, 顾峰雪. 松嫩退化草地芦苇不同叶位叶片营养元素代谢特征[J]. 植物生态学报, 2018, 42(7): 734-740. |
| [10] | 彭曦, 闫文德, 王凤琪, 王光军, 玉昉永, 赵梅芳. 基于叶干质量比的杉木比叶面积估算模型的构建[J]. 植物生态学报, 2018, 42(2): 209-219. |
| [11] | 高景, 王金牛, 徐波, 谢雨, 贺俊东, 吴彦. 不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究[J]. 植物生态学报, 2016, 40(8): 775-787. |
| [12] | 刘波, 吕宪国, 姜明, 张文广, 武海涛. 光照、水深交互作用对松嫩湿地芦苇种子萌发的影响[J]. 植物生态学报, 2015, 39(6): 616-620. |
| [13] | 陈清, 王义东, 郭长城, 王中良. 天津沼泽湿地芦苇叶片的碳稳定同位素比值分布特征及其环境影响因素[J]. 植物生态学报, 2015, 39(11): 1044-1052. |
| [14] | 胡楚琦, 刘金珂, 王天弘, 王文琳, 卢山, 周长芳. 三种盐胁迫对互花米草和芦苇光合作用的影响[J]. 植物生态学报, 2015, 39(1): 92-103. |
| [15] | 张超, 占东霞, 张鹏鹏, 张亚黎, 罗宏海, 张旺锋. 棉花苞叶光呼吸和PSII热耗散对土壤水分的响应[J]. 植物生态学报, 2014, 38(4): 387-395. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19