
植物生态学报 ›› 2025, Vol. 49 ›› Issue (1): 74-82.DOI: 10.17521/cjpe.2024.0095 cstr: 32100.14.cjpe.2024.0095
所属专题: 退化草地恢复理论与技术; 草原与草业
张辉, 赵赟鹏, 刘晓琛, 郭增鹏, 胡国瑞, 冯彦皓, 马妙君*( )(
)( )
)
收稿日期:2024-04-02
接受日期:2024-10-24
出版日期:2025-01-20
发布日期:2025-03-08
通讯作者:
* 马妙君: ORCID: 0000-0001-7046-9574 (mjma@lzu.edu.cn)基金资助:
ZHANG Hui, ZHAO Yun-Peng, LIU Xiao-Chen, GUO Zeng-Peng, HU Guo-Rui, FENG Yan-Hao, MA Miao-Jun*()()
Received:2024-04-02
Accepted:2024-10-24
Online:2025-01-20
Published:2025-03-08
Supported by:摘要:
青藏高原东部高寒草甸局部地区因长期过度放牧而发生退化。土壤种子库是退化生态系统恢复的潜在重要资源, 其在高寒草甸退化过程中的动态及其在退化草地恢复中的作用仍有待探究。该研究在青藏高原东部选择了4个不同退化水平的高寒草甸, 构建了一个退化序列。在这一序列上对植物群落和土壤种子库进行调查, 探究了植物群落及种子库组成和特征在退化序列上的变化规律, 并分析了种子库在植物群落更新和退化高寒草甸恢复中的潜在作用。研究发现: (1)植物群落的物种丰富度和相对多度随高寒草甸的退化而显著降低, 而土壤种子库的物种丰富度呈现先升高后降低的变化规律, 种子密度则显著降低; (2)植物群落与土壤种子库物种组成相似性随退化水平增加而逐渐增加, 即种子库在植物群落更新中的潜在作用随退化水平的增加而增加; (3)相比于植物群落, 种子库对退化的响应有明显的滞后性, 可以在一定程度上缓冲退化对地上植物群落的影响。以上结果说明, 高寒草甸植物群落和土壤种子库对退化的响应不一致, 土壤种子库是退化高寒草甸恢复的重要潜在资源。然而, 土壤种子库对退化的恢复潜力是有限的, 在重度退化水平下, 由于种子库资源的过度损耗, 种子库在修复中的作用下降。以上研究结果可为青藏高原退化高寒草甸的保护、恢复和管理提供科学依据。
张辉, 赵赟鹏, 刘晓琛, 郭增鹏, 胡国瑞, 冯彦皓, 马妙君. 高寒草甸退化过程中土壤种子库的变化及其在植物群落更新中的潜在作用. 植物生态学报, 2025, 49(1): 74-82. DOI: 10.17521/cjpe.2024.0095
ZHANG Hui, ZHAO Yun-Peng, LIU Xiao-Chen, GUO Zeng-Peng, HU Guo-Rui, FENG Yan-Hao, MA Miao-Jun. Dynamics of soil seed bank and its role in plant community regeneration during alpine meadow degradation. Chinese Journal of Plant Ecology, 2025, 49(1): 74-82. DOI: 10.17521/cjpe.2024.0095
| 退化水平 Degradation level | 优势物种 Dominate species | 生境描述 Habitat description |
|---|---|---|
| 对照 Control (I) | 禾叶嵩草、线叶嵩草、垂穗披碱草、草玉梅 Carex hughii, Carex capillifolia, Elymus nutans, and Anemone rivularis | 生长季适度放牧(5-10月), 放牧家畜为牦牛和藏羊, 该样地是青藏高原东部地区典型的高寒草甸。植物群落的盖度为100%。 Moderate grazing in the growing season (from May to October), the grazing animals are yaks (Bos mutus) and Tibetan sheep (Ovis aries). This sample is a typical alpine meadow in the eastern Qingzang Plateau. The plant community coverage was 100%. |
| 轻度退化草甸 Light degraded meadow (II) | 禾叶嵩草、垂穗披碱草、草玉梅、鹅绒委陵菜 Carex hughii, Elymus nutans, Anemone rivularis, and Argentina anserina | 生长季放牧强度过高(5-10月), 放牧强度高于对照, 但低于重度退化草甸样地, 放牧的家畜为牦牛和藏羊, 植物群落出现轻度退化, 群落的优势功能群(莎草科和禾本科)的比例减少, 双子叶杂类草数量更多, 局部地区出现小面积斑块。植物群落的盖度为70%-80%。 Overgrazing in the growing season (from May to October). The grazing intensity was higher than that of the control, and the grazing intensity was lower than that of the moderately degraded meadow. The grazing livestock were yaks and Tibetan sheep, the plant community was slightly degraded, the proportion of dominant functional groups (Cyperaceae and Poaceae) of the alpine meadow community decreased, the number of dicotyledonous weeds increased, and a small gap appeared in local areas. The plant community coverage is 70%-80%. |
| 中度退化草甸 Moderate degraded meadow (III) | 鹅绒委陵菜、草玉梅、密花香薷、细叶亚菊 Argentina anserina, Nemone rivularis, Elsholtzia densa, and Ajania tenuifolia | 生长季过度放牧(5-10月), 放牧强度高于对照和轻度退化草甸, 但低于重度退化草甸样地, 放牧的家畜为牦牛和藏羊, 植物群落出现大面积退化, 群落的优势功能群从莎草科和禾本科转变为双子叶杂类草, 局部地区出现大面积斑块。植物群落的盖度为40%-50%。 Overgrazing in the growing season (from May to October), the grazing intensity was higher than that in the control meadow and light degraded meadows, but lower than that in the seriously degraded meadow plots. The grazing livestock were yaks and Tibetan sheep, and the plant community deteriorated in a large area. The dominant functional group of the community changed from sedge and grass to dicotyledonous weeds, and large gaps appeared in local areas. The plant community coverage is 40%-50%. |
| 重度退化草甸 Seriously degraded meadow (IV) | 密花香薷、鹅绒委陵菜、草玉梅 Elsholtzia densa, Argentina anserina, and Anemone rivularis | 全面过度放牧(0-12月), 全年长期受到牲畜(如牦牛和藏羊)的过度放牧和践踏, 植物群落物种多样性丧失, 几乎没有禾本科和莎草科植物, 群落被双子叶的杂类草占绝对优势。植物群落的盖度为20%-30%。 Overall overgrazing throughout the year (0-12 months), long-term overgrazing and trampling by livestock (yaks and Tibetan sheep), loss of plant community species diversity, almost no grasses and sedges, the community is dominated by dicotyledonous weeds. The plant community coverage is only 20%-30%. |
表1 青藏高原东部高寒草甸4个退化水平下植物群落优势物种、放牧历史和生境描述
Table 1 Description of dominant species of plant communities, grazing history, and habitats under four degradation levels in an alpine meadow on the Qingzang Plateau
| 退化水平 Degradation level | 优势物种 Dominate species | 生境描述 Habitat description |
|---|---|---|
| 对照 Control (I) | 禾叶嵩草、线叶嵩草、垂穗披碱草、草玉梅 Carex hughii, Carex capillifolia, Elymus nutans, and Anemone rivularis | 生长季适度放牧(5-10月), 放牧家畜为牦牛和藏羊, 该样地是青藏高原东部地区典型的高寒草甸。植物群落的盖度为100%。 Moderate grazing in the growing season (from May to October), the grazing animals are yaks (Bos mutus) and Tibetan sheep (Ovis aries). This sample is a typical alpine meadow in the eastern Qingzang Plateau. The plant community coverage was 100%. |
| 轻度退化草甸 Light degraded meadow (II) | 禾叶嵩草、垂穗披碱草、草玉梅、鹅绒委陵菜 Carex hughii, Elymus nutans, Anemone rivularis, and Argentina anserina | 生长季放牧强度过高(5-10月), 放牧强度高于对照, 但低于重度退化草甸样地, 放牧的家畜为牦牛和藏羊, 植物群落出现轻度退化, 群落的优势功能群(莎草科和禾本科)的比例减少, 双子叶杂类草数量更多, 局部地区出现小面积斑块。植物群落的盖度为70%-80%。 Overgrazing in the growing season (from May to October). The grazing intensity was higher than that of the control, and the grazing intensity was lower than that of the moderately degraded meadow. The grazing livestock were yaks and Tibetan sheep, the plant community was slightly degraded, the proportion of dominant functional groups (Cyperaceae and Poaceae) of the alpine meadow community decreased, the number of dicotyledonous weeds increased, and a small gap appeared in local areas. The plant community coverage is 70%-80%. |
| 中度退化草甸 Moderate degraded meadow (III) | 鹅绒委陵菜、草玉梅、密花香薷、细叶亚菊 Argentina anserina, Nemone rivularis, Elsholtzia densa, and Ajania tenuifolia | 生长季过度放牧(5-10月), 放牧强度高于对照和轻度退化草甸, 但低于重度退化草甸样地, 放牧的家畜为牦牛和藏羊, 植物群落出现大面积退化, 群落的优势功能群从莎草科和禾本科转变为双子叶杂类草, 局部地区出现大面积斑块。植物群落的盖度为40%-50%。 Overgrazing in the growing season (from May to October), the grazing intensity was higher than that in the control meadow and light degraded meadows, but lower than that in the seriously degraded meadow plots. The grazing livestock were yaks and Tibetan sheep, and the plant community deteriorated in a large area. The dominant functional group of the community changed from sedge and grass to dicotyledonous weeds, and large gaps appeared in local areas. The plant community coverage is 40%-50%. |
| 重度退化草甸 Seriously degraded meadow (IV) | 密花香薷、鹅绒委陵菜、草玉梅 Elsholtzia densa, Argentina anserina, and Anemone rivularis | 全面过度放牧(0-12月), 全年长期受到牲畜(如牦牛和藏羊)的过度放牧和践踏, 植物群落物种多样性丧失, 几乎没有禾本科和莎草科植物, 群落被双子叶的杂类草占绝对优势。植物群落的盖度为20%-30%。 Overall overgrazing throughout the year (0-12 months), long-term overgrazing and trampling by livestock (yaks and Tibetan sheep), loss of plant community species diversity, almost no grasses and sedges, the community is dominated by dicotyledonous weeds. The plant community coverage is only 20%-30%. |
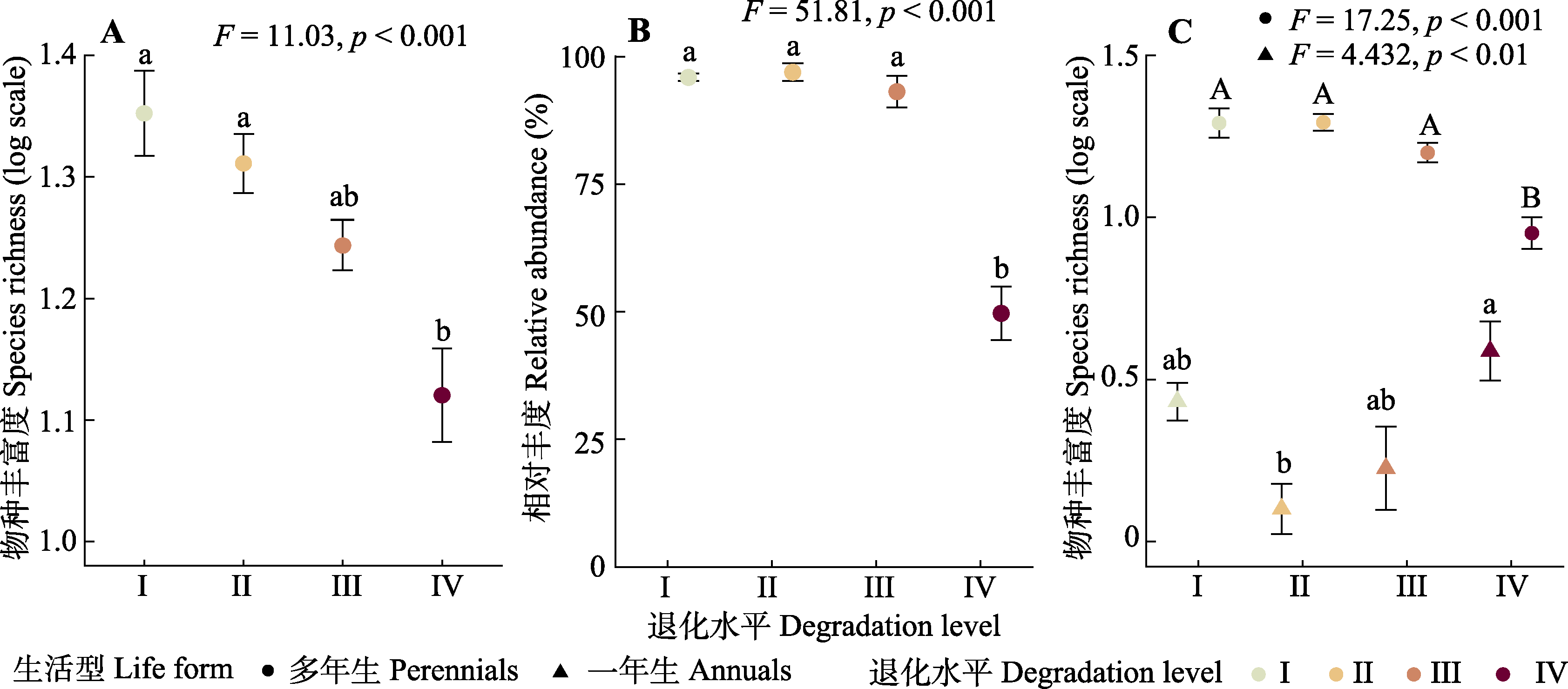
图1 青藏高原东部高寒草甸植物群落物种丰富度(A)、相对丰度(B), 以及不同生活型(C, 一年生和多年生物种)的物种丰富度在4个退化水平之间的变化(平均值±标准误)。I、II、III和IV代表高寒草甸的4个退化水平(对照、轻度退化、中度退化以及重度退化)。不同字母表示不同退化水平间差异显著(p < 0.05)。
Fig. 1 Changes in alpine meadow plant community of species richness (A), relative abundance (B), and species richness of life form (C, annuals and perennials) along four degradation levels (mean ± SE) on the eastern Qingzang Plateau. I to IV represent four degradation levels (control, light degraded meadow, moderate degraded meadow, and seriously degraded meadow). Different letters indicate significant difference between different degradation levels (p < 0.05).
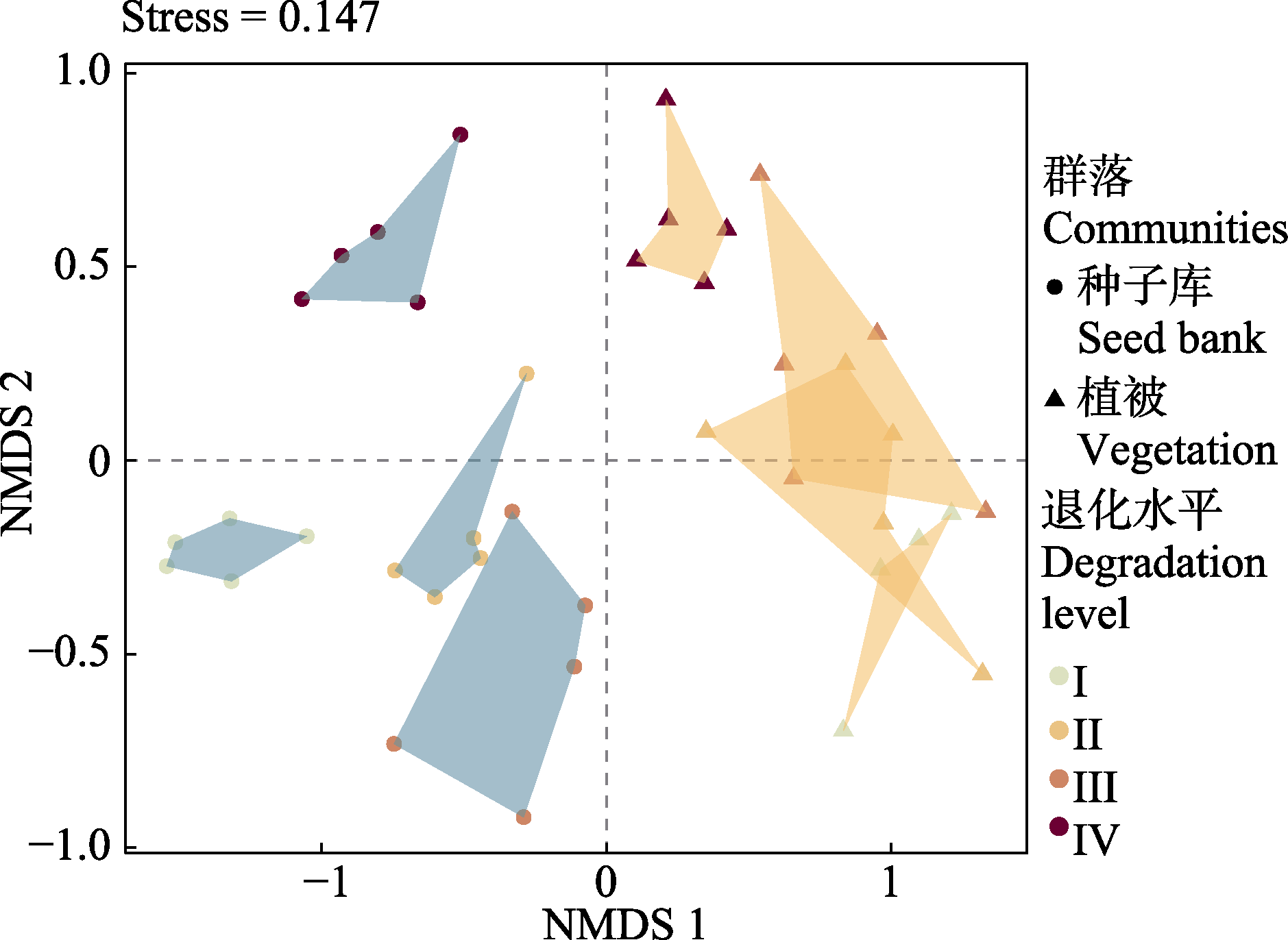
图2 青藏高原东部高寒草甸地上植物群落和土壤种子库的物种组成沿退化梯度的非度量多维尺度分析(NMDS)排序。I、II、III和IV沿颜色由浅到深代表4个退化水平(对照、轻度退化、中度退化以及重度退化)。
Fig. 2 Non-metric Multidimensional Scaling (NMDS) ordination of species composition of the aboveground vegetation and soil seed bank along a degradation gradient in an alpine meadow on the eastern Qingzang Plateau. I to IV represent four degradation levels (control, light degraded meadow, moderate degraded meadow, and seriously degraded meadow) along the color from light to dark.
| 群落类型 Community type | 退化水平 Degradation level | R2 | p |
|---|---|---|---|
| 植物群落 Plant community | I vs II | 0.21 | 0.096 |
| I vs III | 0.25 | 0.046* | |
| I vs IV | 0.57 | 0.015* | |
| II vs III | 0.12 | 0.380 | |
| II vs IV | 0.42 | 0.015* | |
| III vs IV | 0.38 | 0.015* | |
| 土壤种子库 Soil seed bank | I vs II | 0.55 | 0.015* |
| I vs III | 0.62 | 0.015* | |
| I vs IV | 0.57 | 0.015* | |
| II vs III | 0.18 | 0.096 | |
| II vs IV | 0.44 | 0.015* | |
| III vs IV | 0.50 | 0.015* | |
| 植物群落与土壤种子库 Plant community-soil seed bank | I vs I | 0.75 | 0.016* |
| II vs II | 0.46 | 0.016* | |
| III vs III | 0.44 | 0.015* | |
| IV vs IV | 0.49 | 0.015* |
表2 青藏高原东部高寒草甸土壤种子库之间、地上植物群落之间以及植物群落与土壤种子库间的物种组成在4个退化水平之间的差异分析
Table 2 Pairwise comparisons of species using PERMANOVA (Permutational multivariate analysis of variance) based on Bray-Curtis dissimilarity among soil seed banks, plant communities, and species composition between aboveground vegetation and seed bank along a degradation gradient in an alpine meadow on the eastern Qingzang Plateau
| 群落类型 Community type | 退化水平 Degradation level | R2 | p |
|---|---|---|---|
| 植物群落 Plant community | I vs II | 0.21 | 0.096 |
| I vs III | 0.25 | 0.046* | |
| I vs IV | 0.57 | 0.015* | |
| II vs III | 0.12 | 0.380 | |
| II vs IV | 0.42 | 0.015* | |
| III vs IV | 0.38 | 0.015* | |
| 土壤种子库 Soil seed bank | I vs II | 0.55 | 0.015* |
| I vs III | 0.62 | 0.015* | |
| I vs IV | 0.57 | 0.015* | |
| II vs III | 0.18 | 0.096 | |
| II vs IV | 0.44 | 0.015* | |
| III vs IV | 0.50 | 0.015* | |
| 植物群落与土壤种子库 Plant community-soil seed bank | I vs I | 0.75 | 0.016* |
| II vs II | 0.46 | 0.016* | |
| III vs III | 0.44 | 0.015* | |
| IV vs IV | 0.49 | 0.015* |
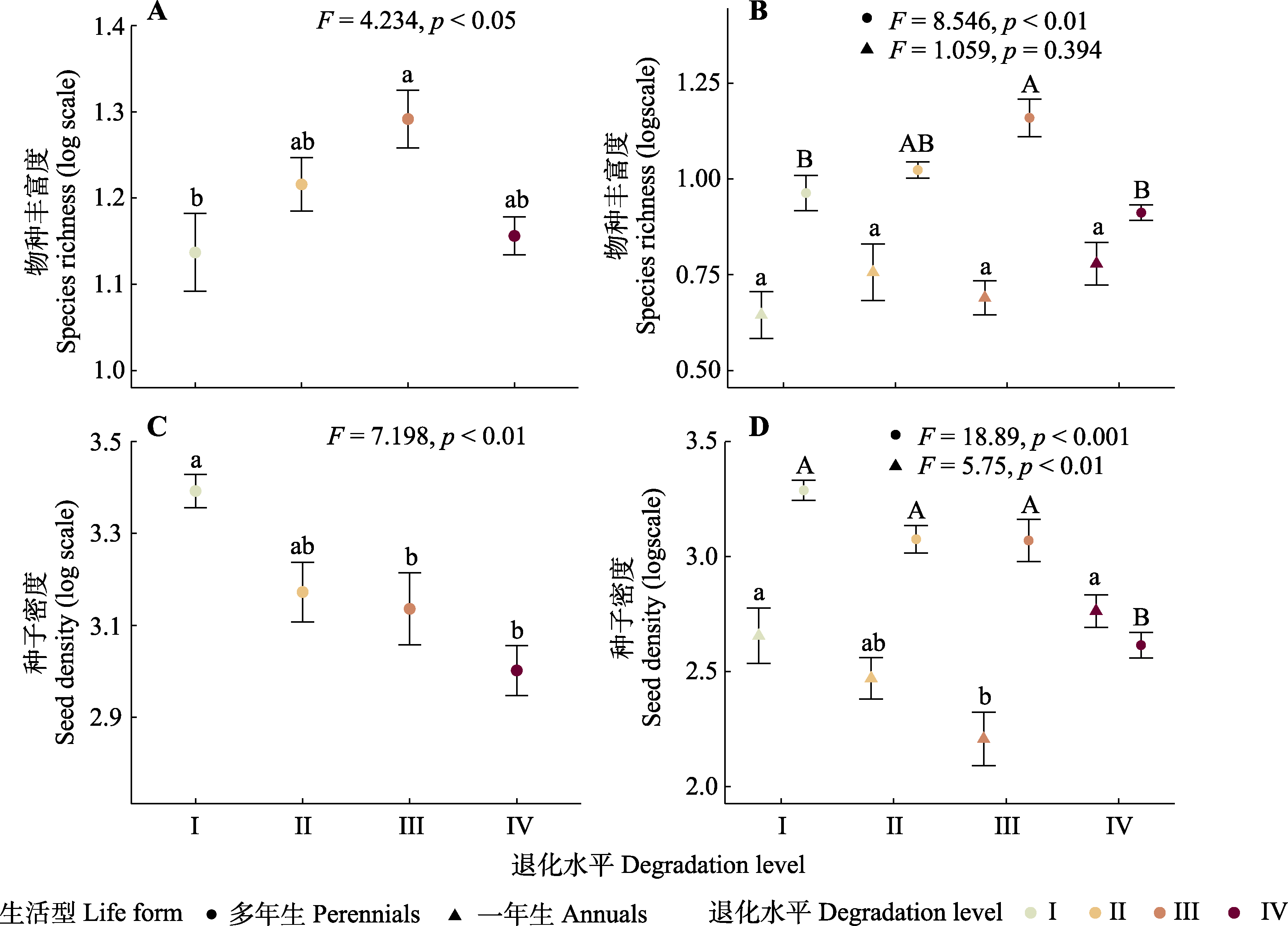
图3 青藏高原东部高寒草甸土壤种子库物种丰富度(A)、不同生活型的物种丰富度(B)、种子密度(C)以及不同生活型的种子密度(D)在4个退化水平之间的变化(平均值±标准误)。I、II、III、和IV代表4个退化水平(对照、轻度退化、中度退化以及重度退化)。不同字母表示不同退化水平间的差异显著(p < 0.05)。
Fig. 3 Changes in species richness (A), species richness of different lifeforms (B), seed density (C), and seed density of different lifeforms (D) in alpine meadow seed bank along four degradation levels (mean ± SE) on the eastern Qingzang Plateau. I to IV represent four degradation levels (control, light degraded meadow, moderate degraded meadow, and seriously degraded meadow). Different letters indicate significant differences among the four degradation levels (p < 0.05).

图4 青藏高原东部高寒草甸土壤种子库和地上植物群落物种组成间的Bray-Curtis不相似性沿退化梯度上的变化趋势。I、II、III和IV代表4个退化水平(对照、轻度退化、中度退化以及重度退化)。
Fig. 4 Bray-Curtis dissimilarity of species composition between seed bank and aboveground vegetation change along a degradation gradient in an alpine meadow on the eastern Qingzang Plateau. I to IV represent four degradation levels (control, light degraded meadow, moderate degraded meadow, and seriously degraded meadow).
| [1] | An H, Baskin CC, Ma M (2022). Nonlinear response of the soil seed bank and its role in plant community regeneration with increased grazing disturbance. Journal of Applied Ecology, 59, 2593-2603. |
| [2] |
An H, Zhao YP, Ma MJ (2020). Precipitation controls seed bank size and its role in alpine meadow community regeneration with increasing altitude. Global Change Biology, 26, 5767-5777.
DOI PMID |
| [3] | Bhattachan A, D’Odorico P, Dintwe K, Okin GS, Collins SL (2014). Resilience and recovery potential of duneland vegetation in the southern Kalahari. Ecosphere, 5, 1-14. DOI: 10.1890/es13-00268.1. |
| [4] | Bossuyt B, Honnay O (2008). Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. Journal of Vegetation Science, 19, 875-884. |
| [5] | Brock MA (2011). Persistence of seed banks in Australian temporary wetlands. Freshwater Biology, 56, 1312-1327. |
| [6] | Chu H, Zhang C, Dong Q, Shang Z, Degen AA, Yang X, Yu Y, Yang Z, Zhang Y (2019). The effect of grazing intensity and season on the soil seed bank and its relation with above-ground vegetation on the alpine steppe. Agriculture Ecosystems & Environment, 285, 106622. DOI: 10.1016/j.agee.2019.106622. |
| [7] | Cooper EJ (2006). Reindeer grazing reduces seed and propagule banks in the high Arctic. Canadian Journal of Botany, 84, 1740-1752. |
| [8] | Eskelinen A, Elwood E, Harrison S, Beyen E, Gremer JR (2021). Vulnerability of grassland seed banks to resource-enhancing global changes. Ecology, 102, e03512. DOI: 10.1002/ecy.3512. |
| [9] | Eskelinen A, Jessen MT, Bahamonde HA, Bakker JD, Borer ET, Caldeira MC, Harpole WS, Jia M, Lannes LS, Nogueira C, Olde Venterink H, Peri PL, Porath-Krause AJ, Seabloom EW, Schroeder K, et al. (2023). Herbivory and nutrients shape grassland soil seed banks. Nature Communications, 14, 3949. DOI: 10.1038/s41467-023-39677-x. |
| [10] | Fenner M, Thompson K (2005). The Ecology of Seeds. Cambridge University Press, Cambridge, UK. |
| [11] | Guo Z, Zhao Y, Zhang P, Zhang H, Baskin CC, Zhang T, Chen Y, Hu G, Yang X, Mao H, Zhang Z, Ma M (2024). Rodents mediate the relationship between seed rain, seed bank and plant community with increased grazing disturbance. Ecological Applications, 34, e2984. DOI: 10.1002/eap.2984. |
| [12] | He JS, Liu ZP, Yao T, Sun SC, Lü Z, Hu XW, Cao GM, Wu XW, Li L, Bu HY, Zhu JX (2020). Analysis of the main constraints and restoration techniques of degraded grassland on the Tibetan Plateau. Science and Technology Review, 38(17), 66-80. |
|
[贺金生, 刘志鹏, 姚拓, 孙书存, 吕植, 胡小文, 曹广民, 吴新卫, 李黎, 卜海燕, 朱剑霄 (2020). 青藏高原退化草地恢复的制约因子及修复技术. 科技导报, 38(17), 66-80.]
DOI |
|
| [13] | He M, Xin C, Baskin CC, Li J, Zhao Y, An H, Sheng X, Zhao L, Zhao Y, Ma M (2021). Different response of transient and persistent seed bank of alpine wetland to grazing disturbance on the Tibetan Plateau. Plant and Soil, 459, 93-107. |
| [14] | Hopfensperger KN (2007). A review of similarity between seed bank and standing vegetation across ecosystems. Oikos, 116, 1438-1448. |
| [15] | Kapás RE, Plue J, Kimberley A, Cousins SAO (2020). Grazing livestock increases both vegetation and seed bank diversity in remnant and restored grasslands. Journal of Vegetation Science, 31, 1053-1065. |
| [16] | Klaus VH, Schäfer D, Prati D, Busch V, Hamer U, Hoever CJ, Kleinebecker T, Mertens D, Fischer M, Hölzel N (2018). Effects of mowing, grazing and fertilization on soil seed banks in temperate grasslands in Central Europe. Agriculture, Ecosystems & Environment, 256, 211-217. |
| [17] | Li J, Okin GS, Alvarez L, Epstein H (2007). Quantitative effects of vegetation cover on wind erosion and soil nutrient loss in a desert grassland of southern New Mexico, USA. Biogeochemistry, 85, 317-332. |
| [18] | Liu M, Zhang Z, Sun J, Wang Y, Wang J, Tsunekawa A, Yibeltal M, Xu M, Chen Y (2020). One-year grazing exclusion remarkably restores degraded alpine meadow at Zoige, eastern Tibetan Plateau. Global Ecology and Conservation, 22, e00951. DOI: 10.1016/j.gecco.2020.e00951. |
| [19] | Loydi A (2019). Effects of grazing exclusion on vegetation and seed bank composition in a mesic mountain grassland in Argentina. Plant Ecology & Diversity, 12, 127-138. |
| [20] | Ma M, Baskin CC, Li WJ, Zhao YP, Zhao Y, Zhao L, Chen N, Du GZ (2019). Seed banks trigger ecological resilience in subalpine meadows abandoned after arable farming on the Tibetan Plateau. Ecological Applications, 29, e01959. DOI: 10.1002/eap.1959. |
| [21] | Ma M, Baskin CC, Yu K, Ma Z, Du G (2017). Wetland drying indirectly influences plant community and seed bank diversity through soil pH. Ecological Indicators, 80, 186-195. |
| [22] | Ma M, Baskin CC, Zhao YP, An H (2023). Light controls alpine meadow community assembly during succession by affecting species recruitment from the seed bank. Ecological Applications, 33, e2782. DOI: 10.1002/eap.2782. |
| [23] | Ma M, Collins SL, Du G (2020). Direct and indirect effects of temperature and precipitation on alpine seed banks in the Tibetan Plateau. Ecological Applications, 30, e02096. DOI: 10.1002/eap.2096. |
| [24] | Ma M, Collins SL, Ratajczak Z, Du G (2021). Soil seed banks, alternative stable state theory, and ecosystem resilience. BioScience, 71, 697-707. |
| [25] |
Ma M, Walck JL, Ma Z, Wang L, Du G (2018). Grazing disturbance increases transient but decreases persistent soil seed bank. Ecological Applications, 28, 1020-1031.
DOI PMID |
| [26] | Ma MJ, Zhou XH, Du GZ (2010a). Role of soil seed bank along a disturbance gradient in an alpine meadow on the Tibet Plateau. Flora, 205, 128-134. |
| [27] | Ma MJ, Zhou XH, Wang G, Ma Z, Du GZ (2013). Seasonal dynamics in alpine meadow seed banks along an altitudinal gradient on the Tibetan Plateau. Plant and Soil, 336, 291-302. |
| [28] | Ma MJ, Zhou XH, Wang G, Ma Z, Du GZ (2010b). Seasonal dynamics in alpine meadow seed banks along an altitudinal gradient on the Tibetan Plateau. Plant and Soil, 336, 291-302. |
| [29] | Matus G, Papp M, Tóthmérész B (2005). Impact of management on vegetation dynamics and seed bank formation of inland dune grassland in Hungary. Flora, 200, 296-306. |
| [30] | Noble A, Palmer SM, Glaves DJ, Crowle A, Brown LE, Holden J (2018). Prescribed burning, atmospheric pollution and grazing effects on peatland vegetation composition. Journal of Applied Ecology, 55, 559-569. |
| [31] | Plue J, van Calster H, Auestad I, Basto S, Bekker RM, Bruun HH, Chevalier R, Decocq G, Grandin U, Hermy M, Jacquemyn H, Jakobsson A, Jankowska-Błaszczuk M, Kalamees R, Koch MA, et al. (2021). Buffering effects of soil seed banks on plant community composition in response to land use and climate. Global Ecology and Biogeography, 30, 128-139. |
| [32] | Sun J, Ma BB, Lu XY (2018). Grazing enhances soil nutrient effects: trade-offs between aboveground and belowground biomass in alpine grasslands of the Tibetan Plateau. Land Degradation & Development, 29, 337-348. |
| [33] | Wang X, Ge W, Zhang M, Fernández-Pascual E, Moles A, Saatkamp A, Rosbakh S, Bu H, Panahi P, Ma M (2024). Large and non-spherical seeds are less likely to form a persistent soil seed bank. Proceedings of the Royal Society B: Biological Sciences, 291, 20232764. DOI: 10.1098/rspb.2023.2764. |
| [34] |
Zhang C, Willis CG, Burghardt LT, Qi W, Liu K, de Moura Souza-Filho PR, Ma Z, Du G (2014). The community-level effect of light on germination timing in relation to seed mass: a source of regeneration niche differentiation. New Phytologist, 204, 496-506.
DOI PMID |
| [35] | Zhao YP, Liao JC, Bao XK, Ma MJ (2021). Soil seed bank dynamics are regulated by bird diversity and soil moisture during alpine wetland degradation. Biological Conservation, 263, 109360. DOI: 10.1016/j.biocon.2021.109360. |
| [1] | 张安宁, 肖亚宁, 赵霞, 张淼, 崔瀚文, 陈书燕, 安黎哲. 青藏高原灌木化与氮添加对土壤线虫及功能性状的交互作用[J]. 植物生态学报, 2026, 50(预发表): 0-. |
| [2] | 李燚, 黄欢, 赵艳超, 陈立同. 养分添加对不同海拔高寒草甸群落多样性及组成的影响[J]. 植物生态学报, 2026, 50(预发表): 0-. |
| [3] | 张静, 陈洁, 李艳朋, 盘李军, 许涵, 李意德, 何海生. 南亚热带针阔混交人工林植物生物量比较及其影响因子分析[J]. 植物生态学报, 2026, 50(2): 400-416. |
| [4] | 张法伟, 李红琴, 祝景彬, 樊博, 周华坤, 李英年, 梁乃申. 氮添加和降水改变对高寒草甸生态系统地上与地下碳储的影响[J]. 植物生态学报, 2025, 49(9): 1399-1409. |
| [5] | 陈刚刚, 朱思洁, 郭亮娜, 付芳伟, 刘昱灼, 李江荣. 藏东南色季拉山高山树线乔灌地上-地下养分分配策略[J]. 植物生态学报, 2025, 49(9): 1515-1526. |
| [6] | 郑子仪, 陈江慧, 刘慧颖. 气候变暖提高青藏高原高寒草甸优势物种的根系分泌速率[J]. 植物生态学报, 2025, 49(9): 1363-1373. |
| [7] | 张琳, 陈华阳, 黄振英. 2004-2010年鄂尔多斯沙地草地长期监测样地植物物种组成和群落特征数据集[J]. 植物生态学报, 2025, 49(8): 1263-1270. |
| [8] | 王尧, 王耀彬, 陈子彦, 伊如汉, 白永飞, 赵玉金, 金晶炜. 连续干旱对蒙古高原草地恢复力和抵抗力的影响[J]. 植物生态学报, 2025, 49(7): 1070-1081. |
| [9] | 陈龙, 郭柯, 勾晓华, 赵秀海, 马泓若. 祁连圆柏林群落组成及特征[J]. 植物生态学报, 2025, 49(6): 852-864. |
| [10] | 王世松, 曲孝云, 董劭琼, 李佳鸿, 杨琦, 侯满福, 赵利清, 郭柯, 刘长成, 胥晓. 西藏札达典型荒漠植被类型及群落特征[J]. 植物生态学报, 2025, 49(5): 801-812. |
| [11] | 王娟, 张登山, 肖元明, 裴全帮, 王博, 樊博, 周国英. 长期围封后高寒草原植物根系分泌物特征与环境因子关系[J]. 植物生态学报, 2025, 49(4): 596-609. |
| [12] | 乔沛阳, 顾肖璇, 刘昌鑫, 曹泽宇, 张婷婷, 林晨, 陈钦常, 彭修凡, 陈菲菲, 李华亮, 陈伟, 陈鹭真. 超强台风“摩羯”登陆点海南东寨港红树林受损状况研究[J]. 植物生态学报, 2025, 49(4): 540-551. |
| [13] | 贾慧琳, 倪隆康, 秦佳双, 廖苏慧, 谭羽, 何佳懿, 顾大形. 极端干旱后喀斯特树木水力功能恢复动态及其影响因素[J]. 植物生态学报, 2025, 49(12): 2004-2014. |
| [14] | 刘洋, 刘傲, 宋璇紫, 杨云, 安柏衡, 韩梦姣, 班玛才格尔, 米玛旺堆. 高原鼠兔干扰对高寒草甸主要植物生态位及种间联结的影响[J]. 植物生态学报, 2025, 49(12): 2054-2068. |
| [15] | 杜华栋, 王梦雨, 聂文杰, 孙浩, 车旭曦, 唐勋. 半干旱矿区塌陷地光伏电站建设对植物群落特征的影响[J]. 植物生态学报, 2025, 49(11): 1778-1790. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()