
植物生态学报 ›› 2018, Vol. 42 ›› Issue (8): 818-830.DOI: 10.17521/cjpe.2018.0056
所属专题: 全球变化与生态系统
刁励玮1,2,*,李平2,3,*,刘卫星2,徐姗4,乔春连5,曾辉1,6,刘玲莉2,3,**( )
)
收稿日期:2018-03-07
出版日期:2018-08-20
发布日期:2018-11-03
通讯作者:
刁励玮,李平,刘玲莉
基金资助:
DIAO Li-Wei1,2,*,LI Ping2,3,*,LIU Wei-Xing2,XU Shan4,QIAO Chun-Lian5,ZENG Hui1,6,LIU Ling-Li2,3,**()
Received:2018-03-07
Online:2018-08-20
Published:2018-11-03
Contact:
Li-Wei DIAO,Ping LI,Ling-Li LIU
Supported by:摘要:
生产力是草地生态系统重要的服务功能, 而生物量作为生态系统生产力的主要组成部分, 往往同时受到氮和水分两个因素的限制。在全球变化背景下, 研究草地生态系统生物量对氮沉降增加和降水变化的响应具有重要意义, 但现有研究缺乏对其在大区域空间尺度以及长时间尺度上响应的综合评估和量化。本研究搜集了1990-2017年间发表论文的有关模拟氮沉降及降水变化研究的相关数据, 进行整合分析, 探讨草地生态系统生物量对氮沉降和降水量两个因素的变化在空间和时间尺度上的响应。结果表明: (1)氮添加、增雨处理以及同时增氮增雨处理都能够显著地提高草地生态系统的地上生物量(37%, 41%, 104%)、总生物量(32%, 23%, 60%)和地上地下生物量比(29%, 25%, 46%)。单独增雨显著提高地下生物量(10%), 单独施氮对地下生物量影响不显著, 但同时增雨则能显著提高地下生物量(43%); (2)氮添加和增雨处理对草地生态系统生物量的影响存在明显的空间变异。在温暖性气候区和海洋性气候区的草地生态系统中, 氮添加对地上、总生物量及地上地下生物量比的促进作用更强, 而在寒冷性气候区和温带大陆性气候区的草地生态系统中, 则增雨处理对地下、总生物量的促进作用更强; (3)草地生态系统生物量对氮添加和增雨处理的响应也存在时间格局上的变化, 地下生物量随着氮添加年限的增加有降低的趋势, 地上、总生物量及地上地下生物量比则有增加的趋势。增雨年限的增加对总生物量没有明显的影响, 但持续促进地上生物量和地下生物量, 增加地上地下生物量比, 可见长期增氮、长期增雨对地上生物量的促进作用更明显。
刁励玮,李平,刘卫星,徐姗,乔春连,曾辉,刘玲莉. 草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应. 植物生态学报, 2018, 42(8): 818-830. DOI: 10.17521/cjpe.2018.0056
DIAO Li-Wei,LI Ping,LIU Wei-Xing,XU Shan,QIAO Chun-Lian,ZENG Hui,LIU Ling-Li. Response of plant biomass to nitrogen addition and precipitation increasing under different climate conditions and time scales in grassland. Chinese Journal of Plant Ecology, 2018, 42(8): 818-830. DOI: 10.17521/cjpe.2018.0056
图1 整合分析站点分布图。
Fig. 1 The map of distribution of all the field experiments in this meta-analysis.
| 氮添加 Nitrogen addition | ||||
|---|---|---|---|---|
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 109 | 56 | 59 | 42 |
| 年际 Inter-annual | 169 | 79 | 74 | 37 |
| 增雨处理 Precipitation increasing | ||||
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 77 | 82 | 71 | 51 |
| 年际 Inter-annual | 62 | 90 | 98 | 32 |
| 增氮增雨处理 Nitrogen addition and precipitation increasing | ||||
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 19 | 21 | 17 | 14 |
| 年际 Inter-annual | 21 | 32 | 27 | 11 |
表1 数据库结构及参数
Table 1 Database structure and parameters
| 氮添加 Nitrogen addition | ||||
|---|---|---|---|---|
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 109 | 56 | 59 | 42 |
| 年际 Inter-annual | 169 | 79 | 74 | 37 |
| 增雨处理 Precipitation increasing | ||||
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 77 | 82 | 71 | 51 |
| 年际 Inter-annual | 62 | 90 | 98 | 32 |
| 增氮增雨处理 Nitrogen addition and precipitation increasing | ||||
| 实验数量 Study numbers | 地上生物量 AGB | 地下生物量 BGB | 总生物量 TB | 地上地下生物量比 AGB/BGB |
| 年均 Annual mean | 19 | 21 | 17 | 14 |
| 年际 Inter-annual | 21 | 32 | 27 | 11 |
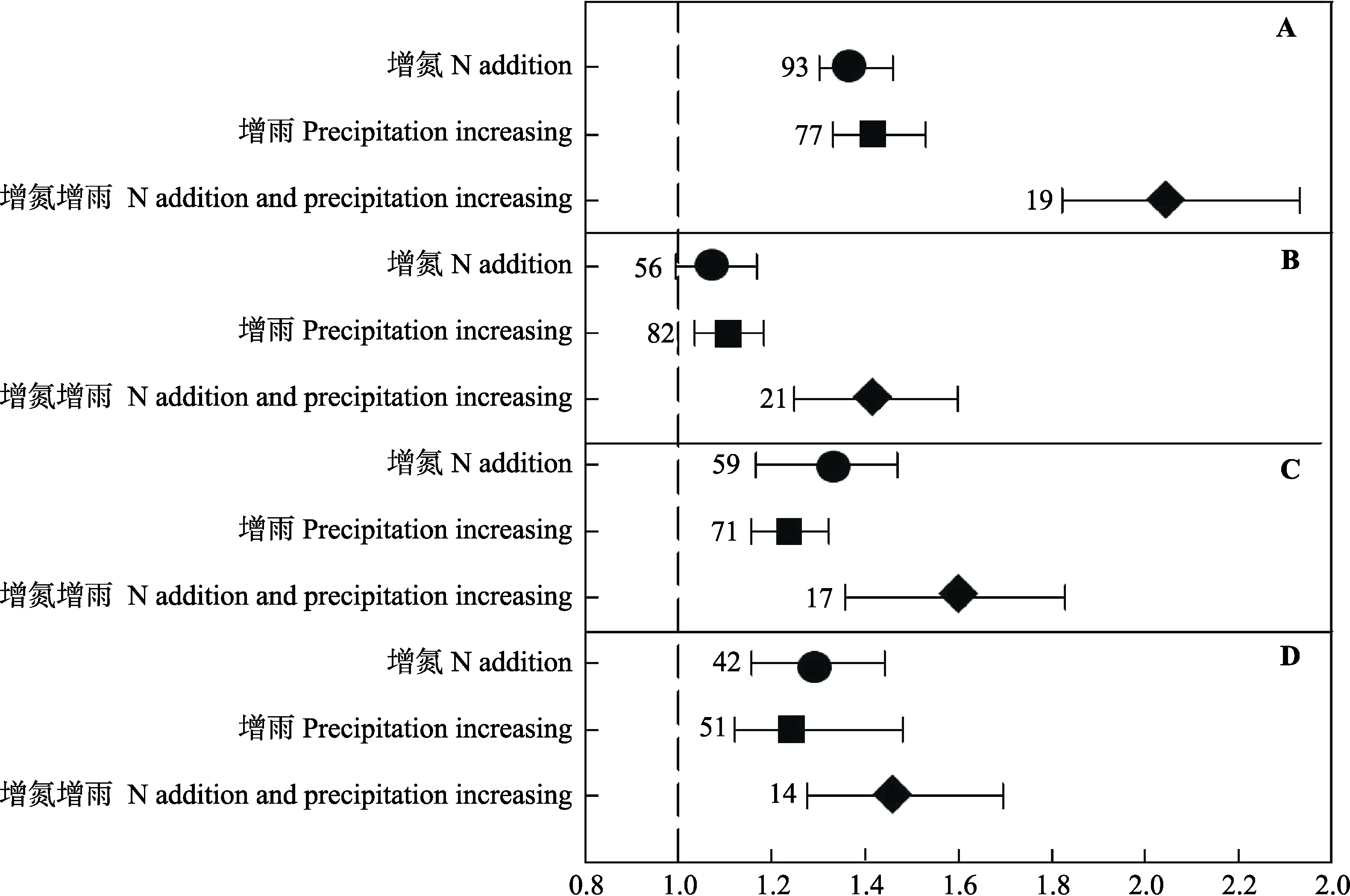
图2 氮添加处理(圆圈)、增雨处理(正方形)及同时增氮增雨处理(菱形)对地上生物量(AGB)(A)、地下生物量(BGB)(B)、总生物量(TB)(C)及地上地下生物量比(AGB/BGB)(D)的影响(平均值± 95%CI)。图中数字代表独立研究数量。
Fig. 2 Effects of N addition (in circle), precipitation increasing (in square) and N addition plus precipitation increasing (in rhombus) on aboveground biomass (AGB)(A), belowground biomass (BGB)(B), total biomass (TB)(C) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D) (mean ± 95%CI). Numbers in the parentheses represent study number.
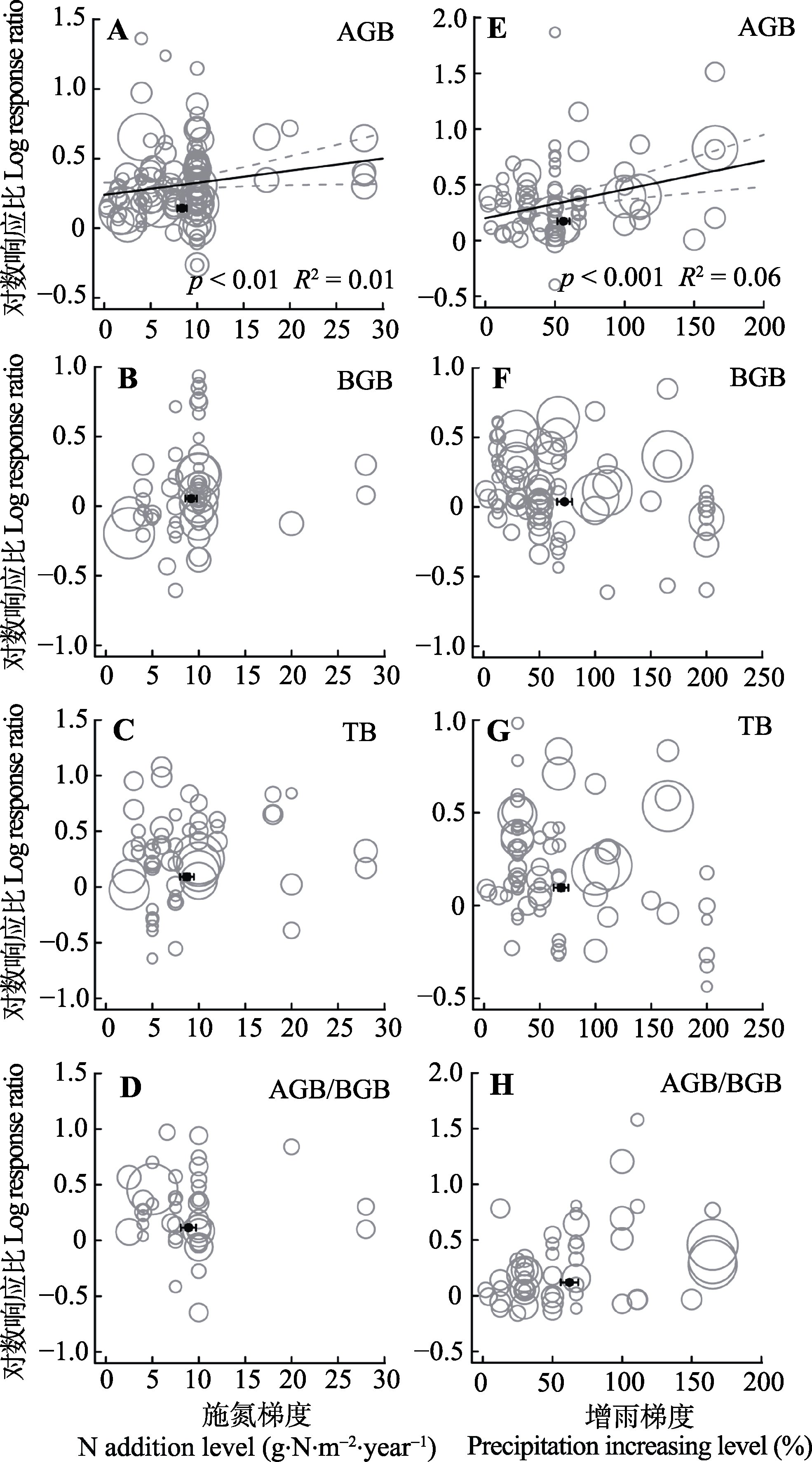
图3 整合回归结果显示地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)沿处理梯度的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度越高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 3 Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass(TB)(C, G) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the treatment level of N addition and precipitation increasing. A-D represent the N addition treatment; E-H represent the precipitation increasing treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta- regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta-regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of treatment level under N addition and precipitation increasing.
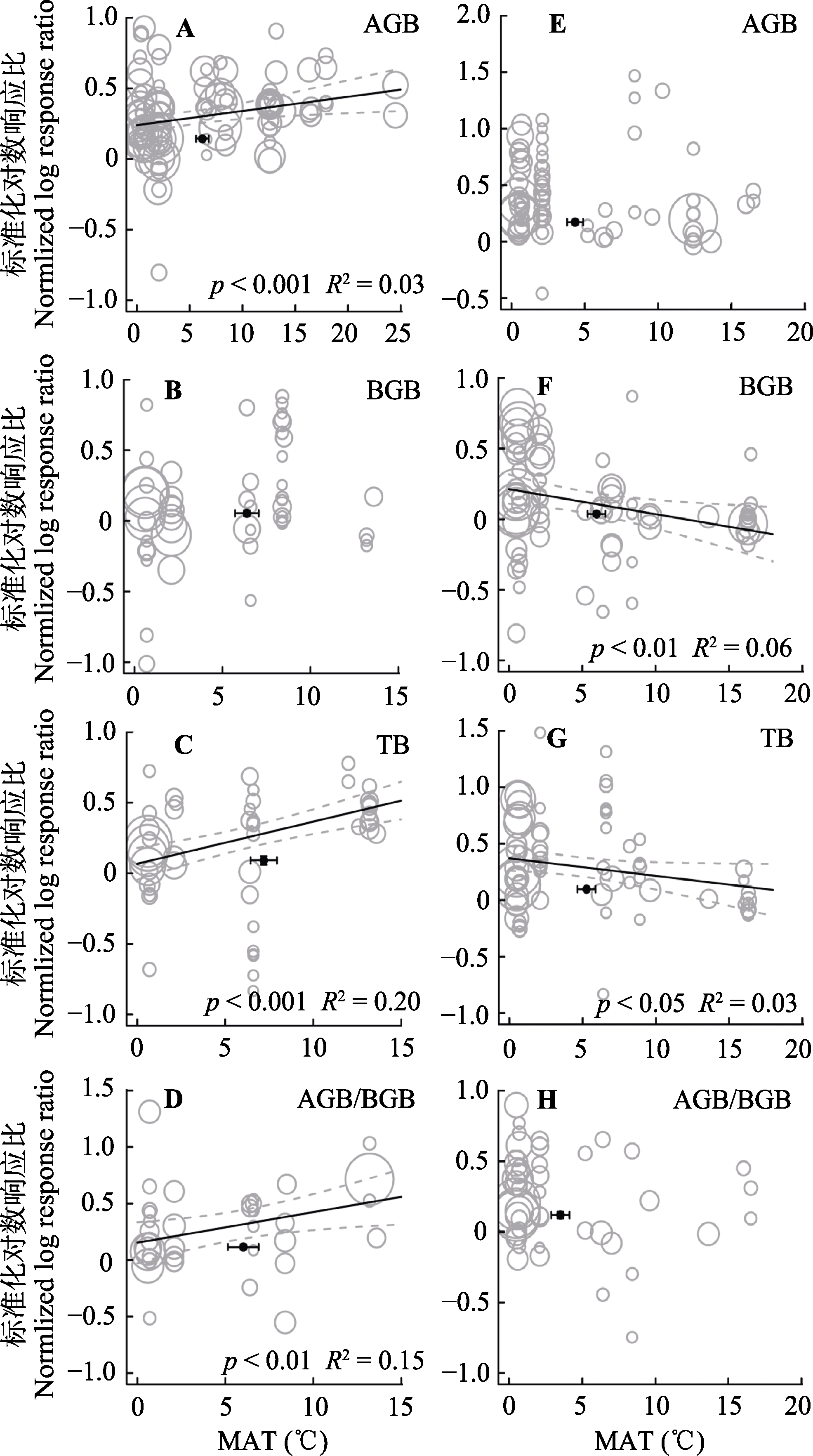
图4 整合回归结果显示随着实验站点年平均气温(MAT)升高, 地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)在不同处理下的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度越高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 4 Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass (TB)(C, G) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the mean annual temperature (MAT) in the study sites. A-D represent the N addition treatment; E-H represent the precipitation increasing treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta-regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta- regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of the means of mean annual temperature (MAT).
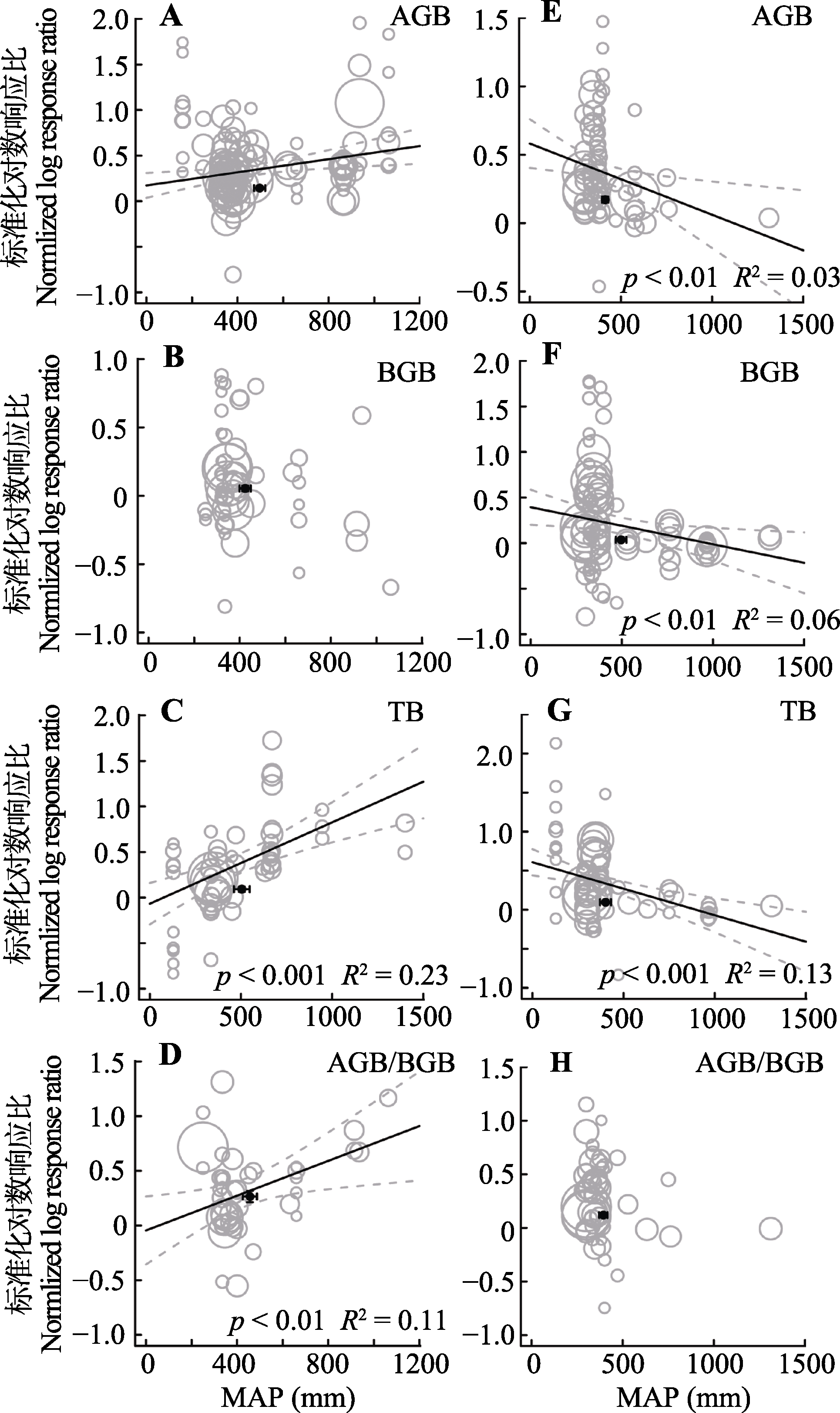
图5 整合回归结果显示随着实验站点年降水量(MAP)增加, 地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)在不同处理下的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度越高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 5 Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass(TB)(C, G) and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the mean annual precipitation (MAP) in the study sites. A-D represent the N addition treatment; E-H represent the precipitation increasing treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta-regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta-regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of the means of mean annual precipitation (MAP).
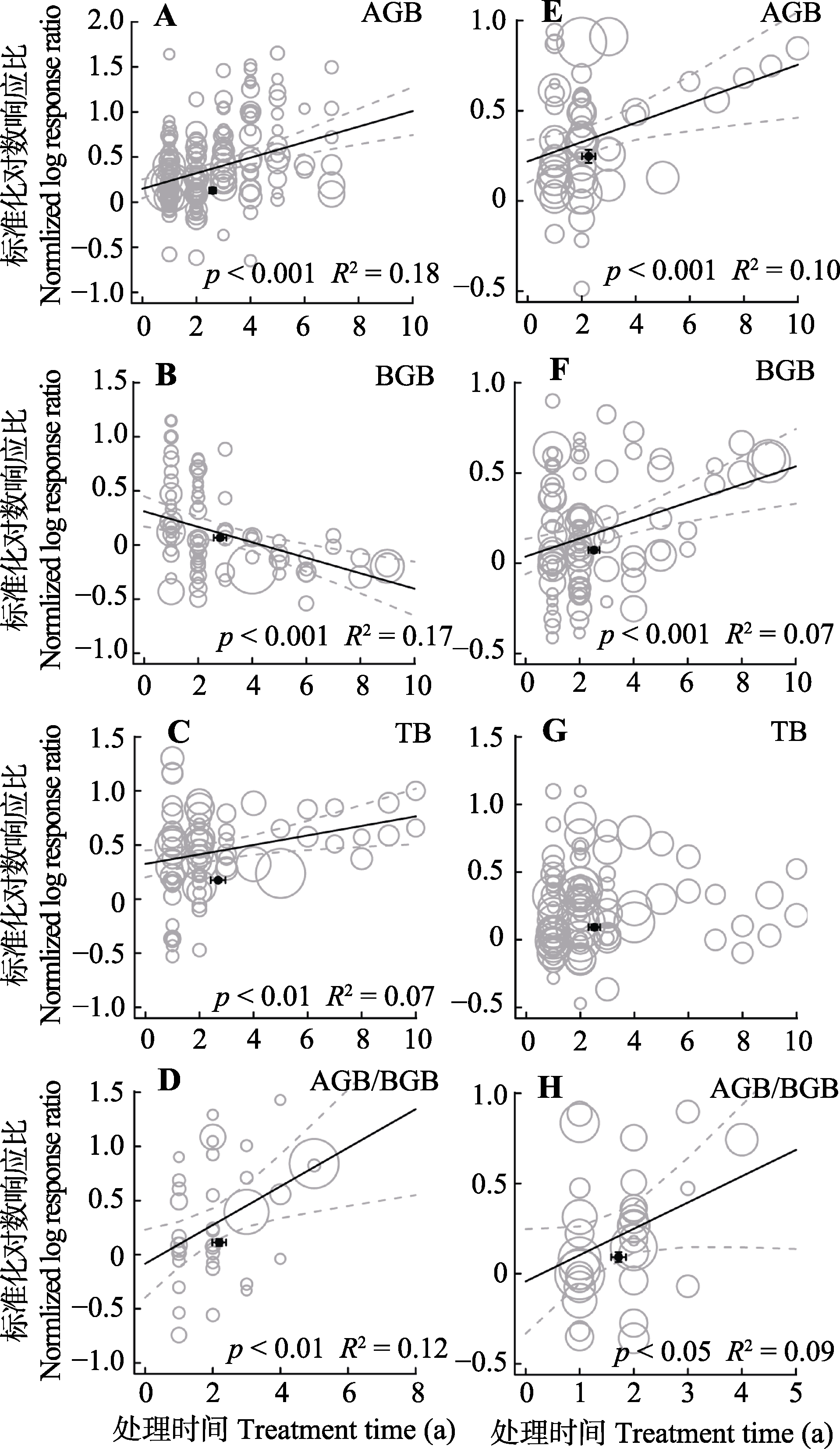
图6 整合回归结果显示随处理年限增加, 地上生物量(AGB)(A, E)、地下生物量(BGB)(B, F)、总生物量(TB)(C, G)和地上地下生物量比(AGB/BGB)(D, H)在不同处理下的变化。A-D代表氮添加, E-H代表增雨处理, 圆圈的大小代表响应比在随机效应meta回归中的权重。圆圈越大, 权重越大, 贡献度高。黑色圆点纵坐标为响应比平均值±标准误差, 横坐标为处理梯度平均值±标准误差。
Fig. 6 Bubble plots of the meta-regression results between the response of aboveground biomass (AGB)(A, E), belowground biomass (BGB)(B, F), total biomass(TB)(C, G)and the ratio of aboveground biomass to belowground biomass (AGB/BGB)(D, H) to the study year. A-D represent the N addition treatment; E-H represent the precipitation increase treatment. The size of the bubble is the relative weight of the effect size (response ratio, RR) in the random-effects meta- regression. Larger bubbles indicate study outcomes that contribute a great overall weight in meta-regression. The y-direction error bars of the black dots represent the standard error of the means of response ratio; the x-direction error bars represent the standard error of the means of treatment time (a).
| [1] |
Adams DC, Gurevitch J, Rosenberg MS ( 1997). Resampling tests for meta-analysis of ecological data. Ecology, 78, 1277-1283.
DOI URL |
| [2] | Bai YF, Li LH, Huang JH, Chen ZZ ( 2001). The influence of plant diversity and functional composition on ecosystem stability of four Stipa communities in the Inner Mongolia Plateau. Acta Botanica Sinica, 43, 280-287. |
| [3] |
Bai YF, Wu JG, Clark CM, Naeem S, Pan QM, Huang JH, Zhang LX, Han XG ( 2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia grasslands. Global Change Biology, 16, 358-372.
DOI URL |
| [4] |
Bai YF, Wu JG, Xing Q, Pan QM, Huang JH, Yang DL, Han XG ( 2008). Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau. Ecology, 89, 2140-2153.
DOI URL PMID |
| [5] |
Bell C, McIntyre N, Cox S, Tissue D, Zak J ( 2008). Soil microbial responses to temporal variations of moisture and temperature in a Chihuahuan Desert grassland. Microbial Ecology, 56, 153-167.
DOI URL PMID |
| [6] |
Bobbink R, Hornung M, Roelofs JG ( 1998). The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. Journal of Ecology, 86, 717-738.
DOI URL |
| [7] |
Brueck H, Erdle K, Gao Y, Giese M, Zhao Y, Peth S, Lin S ( 2010). Effects of N and water supply on water use-efficiency of a semiarid grassland in Inner Mongolia. Plant and Soil, 328, 495-505.
DOI URL |
| [8] |
Burke IC, Lauenroth WK, Parton WJ ( 1997). Regional and temporal variation in net primary production and nitrogen mineralization in grasslands. Ecology, 78, 1330-1340.
DOI URL |
| [9] |
Chen J, Cao JJ, Liu Y, Zhang BC, Wei YL, Ma ZT, Zhu BW ( 2013). Research progress and prospect of nitrogen fertilization on soil respiration. Grassland and Turf, 33(6), 87-93.
DOI URL |
|
[ 陈骥, 曹军骥, 刘玉, 张宝成, 魏永林, 马宗泰, 朱宝文 ( 2013). 氮添加对土壤呼吸影响的研究进展. 草原与草坪, 33(6), 87-93.]
DOI URL |
|
| [10] |
Chen Z, Yu GR, Zhu XJ, Wang QF ( 2014). Spatial pattern and regional characteristics of terrestrial ecosystem carbon fluxes in the northern hemisphere. Quaternary Sciences, 34, 710-722.
DOI URL |
|
[ 陈智, 于贵瑞, 朱先进, 王秋凤 ( 2014). 北半球陆地生态系统碳交换通量的空间格局及其区域特征. 第四纪研究, 34, 710-722.]
DOI URL |
|
| [11] |
Cholaw B, Cubasch U, Yonghui L, Liren J ( 2003). The change of North China climate in transient simulations using the IPCC SRES A2 and B2 scenarios with a coupled atmosphere-ocean general circulation model. Advances in Atmospheric Sciences, 20, 755-766.
DOI URL |
| [12] |
Clark CM, Tilman D ( 2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 451, 712-715.
DOI URL PMID |
| [13] |
Corbin JD, D’Antonio CM ( 2004). Competition between native perennial and exotic annual grasses: Implications for an historical invasion. Ecology, 85, 1273-1283.
DOI URL |
| [14] |
DeLucia EV, Drake JE, Thomas RB, Gonzaiez MI ( 2007). Forest carbon use efficiency: Is respiration a constant fraction of gross primary production? Global Change Biology, 13, 1157-1167.
DOI URL |
| [15] | Dingkuhn M, Luquet D, Clément-Vidal A, Tambour L, Kim HK, Song YH ( 2007). Is plant growth driven by sink regulation? Implications for crop models, phenotyping approaches and ideotypes. Frontis, 155-168. |
| [16] |
Dukes JS, Chiariello NR, Cleland EE, Moore LA, Shaw MR, Thayer S, Field CB ( 2005). Responses of grassland production to single and multiple global environmental changes. PLOS Biology, 3, e319. DOI: 10.1371/journal.pbio.0030319.
DOI URL PMID |
| [17] |
Easterling DR, Meehl GA, Parmesan C, Changnon SA, Karl TR, Mearns LO ( 2000). Climate extremes: Observations, modeling, and impacts. Science, 289, 2068-2074.
DOI URL |
| [18] |
Fang JY, Yu SY, Wu PC, Huang YB, Tsai YH ( 2001). In vitro skin permeation of estradiol from various proniosome formulations.International Journal of Pharmaceutics, 215, 91-99.
DOI URL PMID |
| [19] |
Fang Y, Xun F, Bai WM, Zhang WH, Li LH ( 2012). Long-term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe. PLOS ONE, 7, e47369. DOI: 10.1371/journal.pone.0047369.
DOI URL PMID |
| [20] |
Fay PA, Carlisle JD, Knapp AK, Blair JM, Collins SL ( 2000). Altering rainfall timing and quantity in a mesic grassland ecosystem: Design and performance of rainfall manipulation shelters. Ecosystems, 3, 308-319.
DOI URL |
| [21] |
Fay PA, Prober SM, Harpole WS, Knops JM, Bakker JD, Borer ET, Adler PB ( 2015). Grassland productivity limited by multiple nutrients. Nature Plants, 1, 15080. DOI: 10.1038/nplants.2015.80.
DOI URL PMID |
| [22] |
Gao YZ, Chen Q, Lin S, Giese M, Brueck H ( 2011). Resource manipulation effects on net primary production, biomass allocation and rain-use efficiency of two semiarid grassland sites in Inner Mongolia, China. Oecologia, 165, 855-864.
DOI URL PMID |
| [23] |
Gough L, Shaver GR, Carroll J, Royer DL, Laundre JA ( 2000). Vascular plant species richness in Alaskan arctic tundra: The importance of soil pH. Journal of Ecology, 88, 54-66.
DOI URL |
| [24] |
Goulden ML, McMillan AM, Winston GC, Rocha AV, Manies KL, Harden JW, Bond-Lamberty BP ( 2011). Patterns of NPP, GPP, respiration, and NEP during boreal forest succession. Global Change Biology, 17, 855-871.
DOI URL |
| [25] |
Grechi I, Vivin PH, Hilbert G, Milin S, Robert T, Gaudillere JP ( 2007). Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Environmental and Experimental Botany, 59, 139-149.
DOI URL |
| [26] |
Han BH, Shang ZY, Yuan XB, An Z, Wen HY, Li JB, Fu H, Niu DC ( 2016). Effects of N addition on photosynthetic characteristic and leaf senescence in Stipa bungeana of steppe grasslands in the Loess Plateau. Pratacultural Science, 33, 1070-1076.
DOI URL |
|
[ 韩炳宏, 尚振艳, 袁晓波, 安卓, 文海燕, 李金博, 傅华, 牛得草 ( 2016). 氮添加对黄土高原典型草原长芒草光合特性的影响. 草业科学, 33, 1070-1076.]
DOI URL |
|
| [27] |
Han HG, Shang ZY, Niu DC, Fu H ( 2015). Effect of nitrogen addition on the specific leaf area and important value of dominant species in the typical steppe of the Loess Plateau, China. Acta Agrestia Sinica, 23, 69-74.
DOI URL |
|
[ 韩会阁, 尚振艳, 牛得草, 傅华 ( 2015). 氮添加对黄土高原典型草原植物比叶面积及其重要值的影响. 草地学报, 23, 69-74.]
DOI URL |
|
| [28] |
Harpole WS, Potts DL, Suding KN ( 2007). Ecosystem responses to water and nitrogen amendment in a California grassland. Global Change Biology, 13, 2341-2348.
DOI URL |
| [29] |
Hayes DC, Seastedt TR ( 1987). Root dynamics of tallgrass prairie in wet and dry years. Canadian Journal of Botany, 65, 787-791.
DOI URL |
| [30] |
He M, Dijkstra FA ( 2014). Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytologist, 204, 924-931.
DOI URL |
| [31] |
Hedges LV, Gurevitch J, Curtis PS ( 1999). The meta-analysis of response ratios in experimental ecology. Ecology, 80, 1150-1156.
DOI URL |
| [32] |
Hoekstra JM, Boucher TM, Ricketts TH, Roberts C ( 2005). Confronting a biome crisis: Global disparities of habitat loss and protection. Ecology Letters, 8, 23-29.
DOI URL |
| [33] | Hooper DU, Johnson L ( 1999). Nitrogen limitation in dryland ecosystems: Responses to geographical and temporal variation in precipitation. Biogeochemistry, 46, 247-293. |
| [34] |
Huang WH, Wang SY, Han B, Jiao ZJ, Han GD ( 2014). The research of grassland ecosystem response to simulated atmospheric warming. Pratacultural Science, 31, 2069-2076.
DOI URL |
|
[ 黄文华, 王树彦, 韩冰, 焦志军, 韩国栋 ( 2014). 草地生态系统对模拟大气增温的响应. 草业科学, 31, 2069-2076.]
DOI URL |
|
| [35] | IPCC ( 2012). Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. 582. |
| [36] |
Kang MY, Dai C, Ji WY, Jiang Y, Yuan ZY, Chen HY ( 2013). Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia grasslands, China. PLOS ONE, 8, e69561. DOI: 10.1371/journal.pone.0069561.
DOI URL PMID |
| [37] |
Kelly AE, Goulden ML ( 2008). Rapid shifts in plant distribution with recent climate change. Proceedings of the National Academy of Sciences of the United States of America, 105, 11823-11826.
DOI URL |
| [38] |
Knapp AK, Briggs JM, Koelliker JK ( 2001). Frequency and extent of water limitation to primary production in a mesic temperate grassland. Ecosystems, 4, 19-28.
DOI URL |
| [39] | Koricheva J, Gurevitch J, Mengersen K ( 2013). Handbook of Meta-analysis in Ecology and Evolution. Princeton University Press, Princeton, USA. |
| [40] |
Lauenroth WK, Sala OE ( 1992). Long-term forage production of North American shortgrass steppe. Ecological Applications, 2, 397-403.
DOI URL PMID |
| [41] |
LeBauer DS, Treseder KK ( 2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 89, 371-379.
DOI URL |
| [42] |
Lee M, Manning P, Rist J, Power SA, Marsh C ( 2010). A global comparison of grassland biomass responses to CO2 and nitrogen enrichment. Philosophical Transactions of the Royal Society B: Biological Sciences, 365, 2047-2056.
DOI URL PMID |
| [43] |
Li JZ, Lin S, Taube F, Pan QM, Dittert K ( 2011). Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in Inner Mongolia. Plant and Soil, 340, 253-264.
DOI URL |
| [44] | Li WJ, Liu HM, Zhao JN, Xiu WM, Zhang GL, Huangfu CH, Yang DL ( 2015). Effects of nitrogen and water addition on plant species diversity and biomass of common species in the Stipa baicalensis steppe, Inner Mongolia, China. Acta Ecologica Sinica, 35, 6460-6469. |
| [ 李文娇, 刘红梅, 赵建宁, 修伟明, 张贵龙, 皇甫超河, 杨殿林 ( 2015). 氮和水分添加对贝加尔针茅草原植物多样性及生物量的影响. 生态学报, 35, 6460-6469.] | |
| [45] |
Liu LL, Wang X, Lajeunesse MJ, Miao G, Piao SL, Wan SQ, Deng MF ( 2016). A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes. Global Change Biology, 22, 1394-1405.
DOI URL PMID |
| [46] |
Liu XJ, Zhang Y, Han WX, Tang AH, Shen JL, Cui ZL, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang FS ( 2013). Enhanced nitrogen deposition over China. Nature, 494, 459-462.
DOI URL PMID |
| [47] |
Lu M, Zhou X, Yang Q, Li H, Luo YQ, Fang CM, Chen JK, Yang X, Li B ( 2013). Response of ecosystem carbon cycle to experimental warming: A meta-analysis. Ecology, 94, 726-738.
DOI URL PMID |
| [48] | Lu TT, Zhai XJ, Liu XJ, Wang CJ ( 2014). Influence of short-term enclosure on characteristics of community and species diversity on meadow steppe. Chinese Journal of Grassland, 36(3), 57-60. |
| [ 陆婷婷, 翟夏杰, 刘晓娟, 王成杰 ( 2014). 施肥, 灌溉及火烧对荒漠草原土壤养分和植物群落特征的影响. 中国草地学报, 36(3), 57-60.] | |
| [49] |
Mack MC, Schuur EA, Bret-Harte MS, Shaver GR, Chapin III FS ( 2004). Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature, 431, 440-443.
DOI URL PMID |
| [50] |
Mao W, Li YL, Cui D, Zhao XY, Zhang TH, Li YQ ( 2014). Biomass allocation response of species with different life history strategies to nitrogen and water addition in sandy grassland in Inner Mongolia. Chinese Journal of Plant Ecology, 38, 125-133.
DOI URL |
|
[ 毛伟, 李玉霖, 崔夺, 赵学勇, 张铜会, 李玉强 ( 2014). 沙质草地不同生活史植物的生物量分配对氮和水分添加的响应. 植物生态学报, 38, 125-133.]
DOI URL |
|
| [51] |
Neff JC, Townsend AR, Gleixner G, Lehman SJ, Turnbull J, Bowman WD ( 2002). Variable effects of nitrogen additions on the stability and turnover of soil carbon. Nature, 419, 915-917.
DOI URL PMID |
| [52] |
New M, Lister D, Hulme M, Makin I ( 2002). A high-resolution data set of surface climate over global land areas. Climate Research, 21, 1-25.
DOI URL |
| [53] |
Niu SL, Yang HJ, Zhang Z, Wu MY, Lu Q, Li LH, Wan SQ ( 2009). Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe. Ecosystems, 12, 915-926.
DOI URL |
| [54] |
Pan QM, Bai YF, Wu JG, Han XG ( 2011). Hierarchical plant responses and diversity loss after nitrogen addition: Testing three functionally-based hypotheses in the Inner Mongolia grassland. PLOS ONE, 6, e20078. DOI: 10.1371/journal.pone.0020078.
DOI URL |
| [55] |
Ren HY, Xu ZW, Isbell F, Huang JH, Han XG, Wan SQ, Chen SP, Wang RZ, Zeng DH, Jiang Y, Fang YT ( 2017). Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation. Ecological Monographs, 87, 457-469.
DOI URL |
| [56] |
Rouphael Y, Cardarelli M, Schwarz D, Franken P, Colla G ( 2012). Effects of drought on nutrient uptake and assimilation in vegetable crops. In: Aroca R ed. Plant Responses to Drought Stress. Springer, Berlin. 171-195.
DOI URL |
| [57] |
Sala OE, Parton WJ, Joyce LA, Lauenroth WK ( 1988). Primary production of the central grassland region of the United States. Ecology, 69, 40-45.
DOI URL |
| [58] |
Stevens CJ, Thompson K, Grime JP, Long CJ, Gowing DJ ( 2010). Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition. Functional Ecology, 24, 478-484.
DOI URL |
| [59] | Stocker TF, Qin D, Plattner GK, Tignor M, Allen SK, Boschung J, Midgley PM ( 2013). Climate Change 2013: The Physical Science Basis. Intergovernmental Panel on Climate Change, Working Group I Contribution to the IPCC Fifth Assessment Report (AR5). New York. |
| [60] |
Sugiyama M, Shiogama H, Emori S ( 2010). Precipitation extreme changes exceeding moisture content increases in MIROC and IPCC climate models. Proceedings of the National Academy of Sciences of the United States of America, 107, 571-575.
DOI URL |
| [61] |
Swemmer AM, Knapp AK ( 2008). Defoliation synchronizes aboveground growth of co-occurring C4 grass species. Ecology, 89, 2860-2867.
DOI URL PMID |
| [62] |
Thomey ML, Collins SL, Vargas R, Johnson JE, Brown RF, Natvig DO, Friggens MT ( 2011). Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland. Global Change Biology, 17, 1505-1515.
DOI URL |
| [63] |
Tilman D ( 1985). The resource-ratio hypothesis of plant succession. The American Naturalist, 125, 827-852.
DOI URL |
| [64] |
Tilman D, Wedin D ( 1991). Plant traits and resource reduction for five grasses growing on a nitrogen gradient. Ecology, 72, 685-700.
DOI URL |
| [65] | Viechtbauer W ( 2010). Conducting meta-analyses in R with the metafor package. Journal of Statistic Software, 36, 1-48. |
| [66] | Vitousek PM, Howarth RW ( 1991). Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry, 13, 87-115. |
| [67] |
Wang J, Liu LL, Wang X, Chen YW ( 2015). The interaction between abiotic photodegradation and microbial decomposition under ultraviolet radiation. Global Change Biology, 21, 2095-2104.
DOI URL PMID |
| [68] |
Wang KB, Li JP, Shangguan ZP ( 2012). Biomass components and environmental controls in Ningxia grasslands. Journal of Integrative Agriculture, 11, 2079-2087.
DOI URL |
| [69] | Wang YH, Zhou GS ( 2004). Response of temporal dynamics of aboveground net primary productivity of Leymus chinensis community to precipitation fluctuation in Inner Mongolia. Acta Ecologica Sinica, 24, 1140-1145. |
| [ 王玉辉, 周广胜 ( 2004). 内蒙古羊草草原植物群落地上初级生产力时间动态对降水变化的响应. 生态学报, 24, 1140-1145.] | |
| [70] |
Waring RH, Landsberg JJ, Williams M ( 1998). Net primary production of forests: A constant fraction of gross primary production? Tree Physiology, 18, 129-134.
DOI URL PMID |
| [71] |
Weltzin JF, Loik ME, Schwinning S, Williams DG, Fay PA, Haddad BM, Pockman WT ( 2003). Assessing the response of terrestrial ecosystems to potential changes in precipitation. AIBS Bulletin, 53, 941-952.
DOI URL |
| [72] |
Wrage N, Strodthoff J, Cuchillo HM, Isselstein J, Kayser M ( 2011). Phytodiversity of temperate permanent grasslands: Ecosystem services for agriculture and livestock management for diversity conservation. Biodiversity and Conservation, 20, 3317-3339.
DOI URL |
| [73] |
Wu Z, Dijkstra P, Koch GW, Penuelas J, Hungate BA ( 2011). Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Global Change Biology, 17, 927-942.
DOI URL |
| [74] |
Xia JY, Wan SQ ( 2008). Global response patterns of terrestrial plant species to nitrogen addition. New Phytologist, 179, 428-439.
DOI URL PMID |
| [75] |
Yahdjian L, Gherardi L, Sala OE ( 2011). Nitrogen limitation in arid-subhumid ecosystems: A meta-analysis of fertilization studies. Journal of Arid Environments, 75, 675-680.
DOI URL |
| [76] |
Yan JC, Liang CZ, Fu XY, Wang W, Wang LX, Jia CZ ( 2013). The responses of annual plant traits to rainfall variation in steppe and desert regions. Pratacultural Science, 22(1), 68-76.
DOI URL |
|
[ 闫建成, 梁存柱, 付晓癑, 王炜, 王立新, 贾成朕 ( 2013). 草原与荒漠一年生植物性状对降水变化的响应. 草业学报, 22(1), 68-76.]
DOI URL |
|
| [77] |
Yang YH, Fang JY, Ma WH, Wang W ( 2008). Relationship between variability in aboveground net primary production and precipitation in global grasslands. Geophysical Research Letters, 35(23), 46-63.
DOI URL |
| [78] |
Yuan ZY, Li LH, Han XG, Chen SP, Wang ZW, Chen QS, Bai WM ( 2006). Nitrogen response efficiency increased monotonically with decreasing soil resource availability: A case study from a semiarid grassland in northern China. Oecologia, 148, 564-572.
DOI URL PMID |
| [79] |
Zhou X, Talley M, Luo YQ ( 2009). Biomass, litter, and soil respiration along a precipitation gradient in southern Great Plains, USA. Ecosystems, 12, 1369-1380.
DOI URL |
| [80] |
Zhu K, Chiariello NR, Tobeck T, Fukami T, Field CB ( 2016). Nonlinear, interacting responses to climate limit grassland production under global change. Proceedings of the National Academy of Sciences of the United States of America, 113, 10589-10594.
DOI URL PMID |
| [1] | 黄玲, 王榛, 马泽, 杨发林, 李岚, SEREKPAYEV Nurlan, NOGAYEV Adilbek, 侯扶江. 长期放牧和氮添加对黄土高原典型草原长芒草种群生长的影响[J]. 植物生态学报, 2024, 48(3): 317-330. |
| [2] | 颜辰亦, 龚吉蕊, 张斯琦, 张魏圆, 董学德, 胡宇霞, 杨贵森. 氮添加对内蒙古温带草原土壤活性有机碳的影响[J]. 植物生态学报, 2024, 48(2): 229-241. |
| [3] | 耿雪琪, 唐亚坤, 王丽娜, 邓旭, 张泽凌, 周莹. 氮添加增加中国陆生植物生物量并降低其氮利用效率[J]. 植物生态学报, 2024, 48(2): 147-157. |
| [4] | 舒韦维, 杨坤, 马俊旭, 闵惠琳, 陈琳, 刘士玲, 黄日逸, 明安刚, 明财道, 田祖为. 氮添加对红锥不同序级细根形态和化学性状的影响[J]. 植物生态学报, 2024, 48(1): 103-112. |
| [5] | 赵艳超, 陈立同. 土壤养分对青藏高原高寒草地生物量响应增温的调节作用[J]. 植物生态学报, 2023, 47(8): 1071-1081. |
| [6] | 苏炜, 陈平, 吴婷, 刘岳, 宋雨婷, 刘旭军, 刘菊秀. 氮添加与干季延长对降香黄檀幼苗非结构性碳水化合物、养分与生物量的影响[J]. 植物生态学报, 2023, 47(8): 1094-1104. |
| [7] | 李红琴, 张法伟, 仪律北. 高寒草甸表层土壤和优势植物叶片的化学计量特征对降水改变和氮添加的响应[J]. 植物生态学报, 2023, 47(7): 922-931. |
| [8] | 赵小祥, 朱彬彬, 田秋香, 林巧玲, 陈龙, 刘峰. 叶片凋落物分解的主场优势研究进展[J]. 植物生态学报, 2023, 47(5): 597-607. |
| [9] | 张雅琪, 庞丹波, 陈林, 曹萌豪, 何文强, 李学斌. 荒漠草原土壤氨氧化细菌群落结构对氮添加和枯落物输入的响应[J]. 植物生态学报, 2023, 47(5): 699-712. |
| [10] | 罗来聪, 赖晓琴, 白健, 李爱新, 方海富, Nasir SHAD, 唐明, 胡冬南, 张令. 氮添加背景下土壤真菌和细菌对不同种源入侵植物乌桕生长特征的影响[J]. 植物生态学报, 2023, 47(2): 206-215. |
| [11] | 安凡, 李宝银, 钟全林, 程栋梁, 徐朝斌, 邹宇星, 张雪, 邓兴宇, 林秋燕. 不同种源刨花楠苗木生长与主要功能性状对氮添加的响应[J]. 植物生态学报, 2023, 47(12): 1693-1707. |
| [12] | 葛萍, 李昂, 王银柳, 姜良超, 牛国祥, 哈斯木其尔, 王彦兵, 薛建国, 赵威, 黄建辉. 草甸草原温室气体排放对氮添加量的非线性响应[J]. 植物生态学报, 2023, 47(11): 1483-1492. |
| [13] | 董六文, 任正炜, 张蕊, 谢晨笛, 周小龙. 功能多样性比物种多样性更好解释氮添加对高寒草地生物量的影响[J]. 植物生态学报, 2022, 46(8): 871-881. |
| [14] | 谢欢, 张秋芳, 陈廷廷, 曾泉鑫, 周嘉聪, 吴玥, 林惠瑛, 刘苑苑, 尹云锋, 陈岳民. 氮添加促进丛枝菌根真菌和根系协作维持土壤磷有效性[J]. 植物生态学报, 2022, 46(7): 811-822. |
| [15] | 马炬峰, 辛敏, 徐陈超, 祝琬莹, 毛传澡, 陈欣, 程磊. 丛枝菌根真菌与氮添加对不同根形态基因型水稻氮吸收的影响[J]. 植物生态学报, 2021, 45(7): 728-737. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19