
植物生态学报 ›› 2018, Vol. 42 ›› Issue (9): 955-962.DOI: 10.17521/cjpe.2018.0080
所属专题: 根系生态学
顾伟平1,刘瑞鹏1,李兴欢1,孙涛2,张子嘉2,昝鹏1,温璐宁1,马鹏宇1,毛子军1,*( )
)
收稿日期:2018-04-10
修回日期:2018-09-10
出版日期:2018-09-20
发布日期:2019-01-15
通讯作者:
毛子军
基金资助:
GU Wei-Ping1,LIU Rui-Peng1,LI Xing-Huan1,SUN Tao2,ZHANG Zi-Jia2,ZAN Peng1,WEN Lu-Ning1,MA Peng-Yu1,MAO Zi-Jun1,*()
Received:2018-04-10
Revised:2018-09-10
Online:2018-09-20
Published:2019-01-15
Contact:
Zi-Jun MAO
Supported by:摘要:
细根分解是森林生态系统土壤碳和养分的主要输入途径, 但目前人们对于影响细根分解的主要因素和细根分解模式的了解仍然很少。该研究采用根序划分等级方法, 将红松(Pinus koraiensis)、落叶松(Larix gmelinii)、水曲柳(Fraxinus mandschurica)和白桦(Betula platyphylla)细根组分前四级根划分为两个等级: 一级和二级根混合为低级根, 三级和四级根混合为高级根。利用埋袋法对东北地区4个树种不同根序细根进行连续4年的分解实验, 并对其分解速率以及影响因素进行研究。结果显示, 红松低级根和高级根分解系数分别为0.342和0.461, 落叶松依次分别为0.304和0.436, 水曲柳分别为0.450和0.555, 白桦为0.441和0.579。4个树种均显示低级根分解速率较慢, 而高级根分解速率较快。实验表明, 根系分解系数与酸不溶性物质(AUF)和非结构性碳水化合物(TNC)均具有显著相关性。出现上述结果的主要原因是低级根含有较多的AUF, 很难被分解, 以及含有较少的TNC, 为分解者提供能量较少。
顾伟平, 刘瑞鹏, 李兴欢, 孙涛, 张子嘉, 昝鹏, 温璐宁, 马鹏宇, 毛子军. 四个典型温带树种不同根序细根分解速率及其主要影响因素. 植物生态学报, 2018, 42(9): 955-962. DOI: 10.17521/cjpe.2018.0080
GU Wei-Ping, LIU Rui-Peng, LI Xing-Huan, SUN Tao, ZHANG Zi-Jia, ZAN Peng, WEN Lu-Ning, MA Peng-Yu, MAO Zi-Jun. Decomposition of different root branch orders and its dominant controlling factors in four temperate tree species. Chinese Journal of Plant Ecology, 2018, 42(9): 955-962. DOI: 10.17521/cjpe.2018.0080
| 林型 Forest type | T10 (℃) | W10 (g·g-1) | 林龄 Age (a) | 密度 Density (trees·hm-2) | DBH (cm) | 主要树种 Dominant tree species |
|---|---|---|---|---|---|---|
| 落叶松人工林 Larix gmelinii plantation | 11.6 | 0.396 | 40 | 1 682 | 18.7 | 落叶松 Larix gmelinii |
| 红松人工林 Pinus koraiensis plantation | 10.8 | 0.458 | 46 | 2 904 | 16.1 | 红松 Pinus koraiensis |
| 杨-桦林 Aspen-birch forest | 12.9 | 0.552 | 58 | 2 381 | 12.9 | 山杨、白桦 Populus davidiana, Betula platyphylla |
| 阔叶林 Hardwood forest | 12.3 | 0.579 | 49 | 2 017 | 15.6 | 水曲柳、胡桃楸 Fraxinus mandschurica, Juglans mandshurica |
表1 4种森林类型的林分特征
Table 1 Stand characteristics of the four forest types
| 林型 Forest type | T10 (℃) | W10 (g·g-1) | 林龄 Age (a) | 密度 Density (trees·hm-2) | DBH (cm) | 主要树种 Dominant tree species |
|---|---|---|---|---|---|---|
| 落叶松人工林 Larix gmelinii plantation | 11.6 | 0.396 | 40 | 1 682 | 18.7 | 落叶松 Larix gmelinii |
| 红松人工林 Pinus koraiensis plantation | 10.8 | 0.458 | 46 | 2 904 | 16.1 | 红松 Pinus koraiensis |
| 杨-桦林 Aspen-birch forest | 12.9 | 0.552 | 58 | 2 381 | 12.9 | 山杨、白桦 Populus davidiana, Betula platyphylla |
| 阔叶林 Hardwood forest | 12.3 | 0.579 | 49 | 2 017 | 15.6 | 水曲柳、胡桃楸 Fraxinus mandschurica, Juglans mandshurica |
| 树种 Tree species | 根直径 Root diameter (mm) | |||
|---|---|---|---|---|
| 一级 First-order | 二级 Second-order | 三级 Third-order | 四级 Fourth-order | |
| 红松 Pinus koraiensis | 0.29 ± 0.01 | 0.29 ± 0.02 | 0.53 ± 0.03 | 1.59 ± 0.12 |
| 落叶松 Larix gmelinii | 0.26 ± 0.02 | 0.28 ± 0.01 | 0.46 ± 0.05 | 0.85 ± 0.09 |
| 水曲柳 Fraxinus mandschurica | 0.34 ± 0.01 | 0.45 ± 0.01 | 0.55 ± 0.02 | 0.89 ± 0.07 |
| 白桦 Betula platyphylla | 0.21 ± 0.01 | 0.26 ± 0.03 | 0.27 ± 0.06 | 0.43 ± 0.09 |
表2 4个温带树种一级至四级根平均直径(平均值±标准误差)
Table 2 Mean root diameter from first-order to fourth-order roots of the four temperate tree species (mean ± SE)
| 树种 Tree species | 根直径 Root diameter (mm) | |||
|---|---|---|---|---|
| 一级 First-order | 二级 Second-order | 三级 Third-order | 四级 Fourth-order | |
| 红松 Pinus koraiensis | 0.29 ± 0.01 | 0.29 ± 0.02 | 0.53 ± 0.03 | 1.59 ± 0.12 |
| 落叶松 Larix gmelinii | 0.26 ± 0.02 | 0.28 ± 0.01 | 0.46 ± 0.05 | 0.85 ± 0.09 |
| 水曲柳 Fraxinus mandschurica | 0.34 ± 0.01 | 0.45 ± 0.01 | 0.55 ± 0.02 | 0.89 ± 0.07 |
| 白桦 Betula platyphylla | 0.21 ± 0.01 | 0.26 ± 0.03 | 0.27 ± 0.06 | 0.43 ± 0.09 |
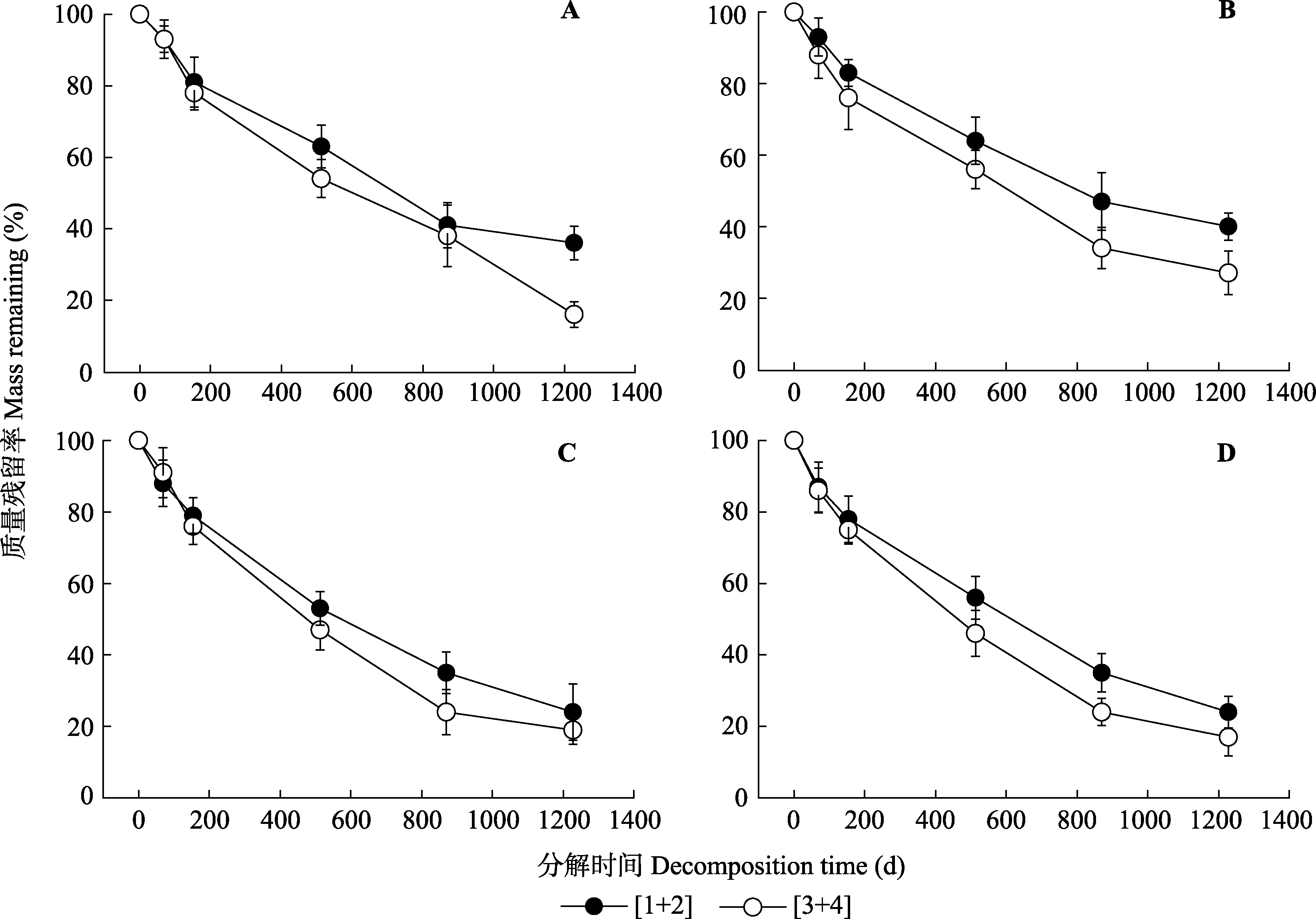
图1 4个树种不同根序不同分解时间的质量残留率(平均值±标准误差)。[1+2]是指1-2级根的合并, [3+4]是指3-4级根的合并。A, 红松。B, 落叶松。C, 水曲柳。D, 白桦。
Fig. 1 Mass remaining (%) of roots of different branch orders of the four tree species (mean ± SE). [1+2] refers to root orders 1 to 2, [3+4] refers to root orders 3 to 4. A, Pinus koraiensis. B, Larix gmelinii. C, Fraxinus mandschurica. D, Betula platyphylla.
| 根序 Root order | 红松 Pinus koraiensis | 落叶松 Larix gmelinii | 水曲柳 Fraxinus mandschurica | 白桦 Betula platyphylla | ||||
|---|---|---|---|---|---|---|---|---|
| k | R2 | k | R2 | k | R2 | k | R2 | |
| [1+2] | 0.342 | 0.983 | 0.304 | 0.985 | 0.450 | 0.993 | 0.441 | 0.987 |
| [3+4] | 0.461 | 0.987 | 0.436 | 0.980 | 0.555 | 0.995 | 0.579 | 0.994 |
表3 每个树种根系的分解常数(k)和相关系数(R2)
Table 3 Decay constants (k) and the correlation coefficient (R2) of root decomposition for each species
| 根序 Root order | 红松 Pinus koraiensis | 落叶松 Larix gmelinii | 水曲柳 Fraxinus mandschurica | 白桦 Betula platyphylla | ||||
|---|---|---|---|---|---|---|---|---|
| k | R2 | k | R2 | k | R2 | k | R2 | |
| [1+2] | 0.342 | 0.983 | 0.304 | 0.985 | 0.450 | 0.993 | 0.441 | 0.987 |
| [3+4] | 0.461 | 0.987 | 0.436 | 0.980 | 0.555 | 0.995 | 0.579 | 0.994 |
| 树种 Tree Species | 红松 Pinus koraiensis | 落叶松 Larix gmelinii | 水曲柳 Fraxinus mandschurica | 白桦 Betula platyphylla | ||||
|---|---|---|---|---|---|---|---|---|
| 根序 Root order | [1+2] | [3+4] | [1+2] | [3+4] | [1+2] | [3+4] | [1+2] | [3+4] |
| N (mg·g-1) | 21.6 ± 0.9 | 12.3 ± 0.5 | 25.5 ± 2.3 | 13.9 ± 1.2 | 31.9 ± 1.8 | 18.3 ± 1.3 | 28.1 ± 1.6 | 16.7 ± 1.1 |
| P (mg·g-1) | 1.9 ± 0.4 | 1.4 ± 0.4 | 2.8 ± 0.2 | 1.9 ± 0.2 | 2.5 ± 0.1 | 1.7 ± 0.2 | 2.2 ± 0.1 | 1.4 ± 0.2 |
| K (mg·g-1) | 4.4 ± 0.5 | 2.9 ± 0.1 | 5.8 ± 0.2 | 4.5 ± 0.3 | 4.0 ± 0.1 | 4.9 ± 0.3 | 5.3 ± 0.6 | 3.7 ± 0.3 |
| AUF (mg·g-1) | 528.5 ± 40.1 | 398.6 ± 32.9 | 513.4 ± 38.7 | 435.8 ± 37.5 | 401.7 ± 29.6 | 289.3 ± 26.4 | 397.5 ± 34.1 | 302.1 ± 28.2 |
| TNC (mg·g-1) | 101.7 ± 10.9 | 150.2 ± 18.4 | 98.3 ± 9.6 | 138.0 ± 14.1 | 195.8 ± 20.6 | 279.1 ± 23.5 | 164.9 ± 18.3 | 214.6 ± 21.4 |
| C:N | 29.6 ± 1.4 | 48.2 ± 1.9 | 23.6 ± 1.8 | 45.2 ± 3.1 | 14.4 ± 2.1 | 23.7 ± 2.4 | 18.7 ± 2.0 | 34.8 ± 3.1 |
表4 四个温带树种的细根分解袋实验的初始化学参数(平均值±标准误差)
Table 4 Initial root chemistry parameters at the start of the litterbag experiment in the four temperate tree species (mean ± SE)
| 树种 Tree Species | 红松 Pinus koraiensis | 落叶松 Larix gmelinii | 水曲柳 Fraxinus mandschurica | 白桦 Betula platyphylla | ||||
|---|---|---|---|---|---|---|---|---|
| 根序 Root order | [1+2] | [3+4] | [1+2] | [3+4] | [1+2] | [3+4] | [1+2] | [3+4] |
| N (mg·g-1) | 21.6 ± 0.9 | 12.3 ± 0.5 | 25.5 ± 2.3 | 13.9 ± 1.2 | 31.9 ± 1.8 | 18.3 ± 1.3 | 28.1 ± 1.6 | 16.7 ± 1.1 |
| P (mg·g-1) | 1.9 ± 0.4 | 1.4 ± 0.4 | 2.8 ± 0.2 | 1.9 ± 0.2 | 2.5 ± 0.1 | 1.7 ± 0.2 | 2.2 ± 0.1 | 1.4 ± 0.2 |
| K (mg·g-1) | 4.4 ± 0.5 | 2.9 ± 0.1 | 5.8 ± 0.2 | 4.5 ± 0.3 | 4.0 ± 0.1 | 4.9 ± 0.3 | 5.3 ± 0.6 | 3.7 ± 0.3 |
| AUF (mg·g-1) | 528.5 ± 40.1 | 398.6 ± 32.9 | 513.4 ± 38.7 | 435.8 ± 37.5 | 401.7 ± 29.6 | 289.3 ± 26.4 | 397.5 ± 34.1 | 302.1 ± 28.2 |
| TNC (mg·g-1) | 101.7 ± 10.9 | 150.2 ± 18.4 | 98.3 ± 9.6 | 138.0 ± 14.1 | 195.8 ± 20.6 | 279.1 ± 23.5 | 164.9 ± 18.3 | 214.6 ± 21.4 |
| C:N | 29.6 ± 1.4 | 48.2 ± 1.9 | 23.6 ± 1.8 | 45.2 ± 3.1 | 14.4 ± 2.1 | 23.7 ± 2.4 | 18.7 ± 2.0 | 34.8 ± 3.1 |
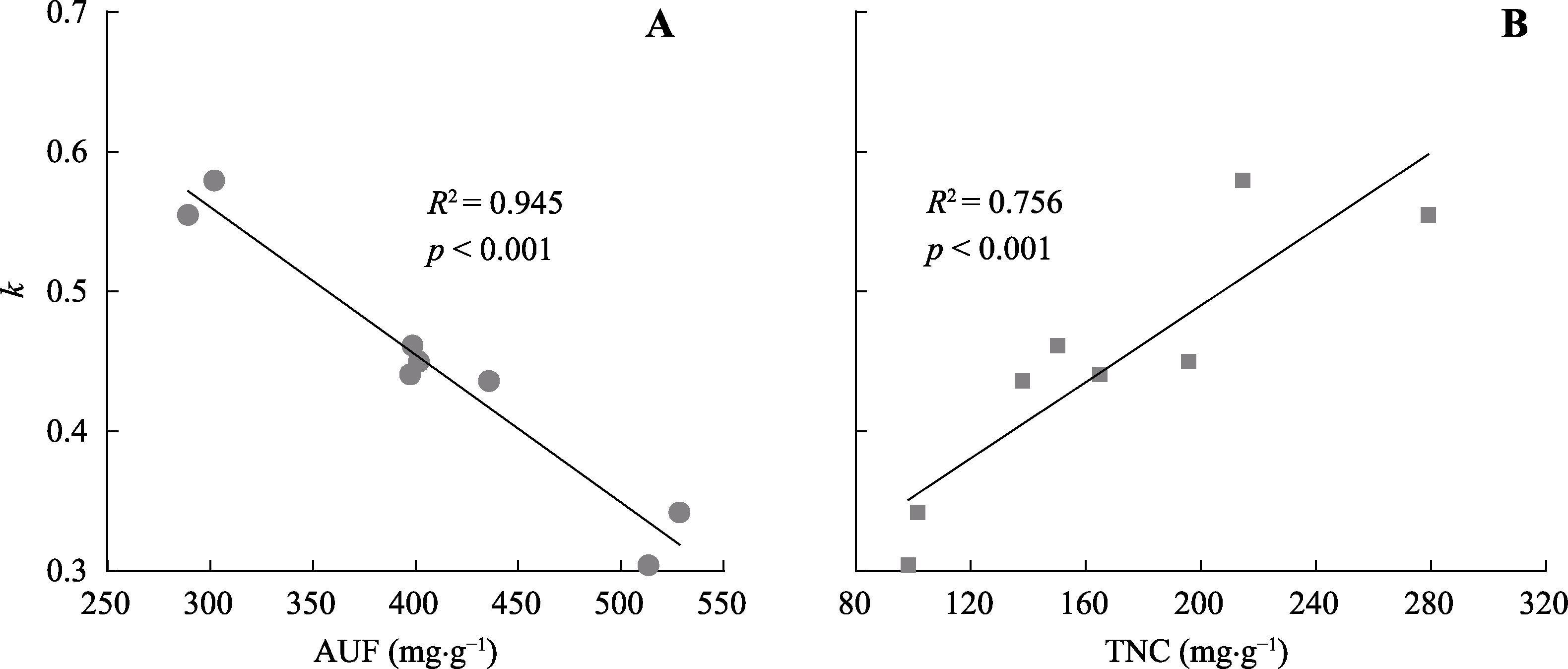
图2 所有树种根系分解常数(k)与酸不溶性物质(AUF)(A)和非结构性碳水化合物(TNC)(B)之间的线性回归关系。
Fig. 2 Regression relation between root decay constants (k) and root initial acid-unhydrolyzable fraction (AUF)(A) and total non-structural carbohydrat (TNC)(B) concentrations across all species.
| [1] |
Aerts R ( 2006). The freezer defrosting: Global warming and litter decomposition rates in cold biomes. Journal of Ecology, 94, 713-724.
DOI URL |
| [2] |
Chen H, Harmon ME, Griffiths RP ( 2001). Decomposition and nitrogen release from decomposition woody roots in coniferous forests of the Pacific Northwest. Canadian Journal of Forest Research, 31, 246-260.
DOI URL |
| [3] |
Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Pérez-Harguindeguy N ( 2008). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecology Letters, 11, 1065-1071.
DOI URL |
| [4] |
Dornbush ME, Isenhart TM, Raich JW ( 2002). Quantifying fine root decomposition: An alternative to buried litterbags. Ecology, 83, 2985-2990.
DOI URL |
| [5] |
Fan P, Guo D ( 2010). Slow decomposition of lower order roots: A key mechanism of root carbon and nutrient retention in the soil. Oecologia, 163, 509-515.
DOI URL PMID |
| [6] |
Goebel M, Hobbie SE, Bulaj B, Zadworny M, Archibald DD, Oleksyn J, Reich PB, Eissenstat DM ( 2011). Decomposition of the finest root branching orders linking belowground dynamics to fine-root function and structure. Ecological Monographs, 81, 89-102.
DOI URL |
| [7] |
Gu JC, Wang DN, Xia XX, Wang SZ ( 2016). Applications of functional classification methods for tree fine root biomass estimation: Advancements and synthesis. Chinese Journal of Plant Ecology, 40, 1344-1351.
DOI URL |
|
[ 谷加存, 王东男, 夏秀雪, 王韶仲 ( 2016). 功能划分方法在树木细根生物量研究中的应用: 进展与评述. 植物生态学报, 40, 1344-1351.]
DOI URL |
|
| [8] |
Gu LC, Wang GL, Jing H, Yao X ( 2017). Response of decomposition and nutrient release in different diameter fine roots of Pinus tabuliformis plantation to N addition. Chinese Journal of Applied Ecology, 28, 2771-2777.
DOI URL |
|
[ 谷利茶, 王国梁, 景航, 姚旭 ( 2017). 氮添加对油松不同径级细根分解及其养分释放的影响. 应用生态学报, 28, 2771-2777.]
DOI URL |
|
| [9] |
Guo D, Mitchell RJ, Withington JM, Fan PP, Hendricks JJ ( 2008a ). Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: Root branch order predominates. Journal of Ecology, 96, 737-745.
DOI URL |
| [10] |
Guo D, Xia M, Wei X, Chang W, Liu Y, Wang Z ( 2008b ). Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytologist, 180, 673-683.
DOI URL PMID |
| [11] |
Guo DL, Mitchell RJ, Hendricks JJ ( 2004). Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest. Oecologia, 140, 450-457.
DOI URL PMID |
| [12] |
Hendricks JJ, Wilson CA, Boring LR ( 2002). Foliar litter position and decomposition in a fire-maintained longleaf pine-wiregrass ecosystem. Canadian Journal of Forest Research, 32, 928-941.
DOI URL |
| [13] |
Hobbie SE, Oleksyn J, Eissenstat DM, Reich PB ( 2010). Fine root decomposition rates do not mirror those of leaf litter among temperate tree species. Oecologia, 162, 505-513.
DOI URL PMID |
| [14] |
Hobbie SE, Reich PB, Ogdahl M, Ogdahl M, Zytkowiak R, Hale C, Karolewski P ( 2006). Tree species on decomposition and forest floor dynamics in a common garden. Ecology, 87, 2288-2297.
DOI URL PMID |
| [15] |
Li H, Wu FZ, Yang WQ, Xu LY, Ni XY, He J, Chang CH ( 2015). Effects of snow cover on acid-soluble extractive and acid-insoluble residue during foliar litter decomposition in the alpine forest. Acta Ecologica Sinica, 35, 4687-4698.
DOI URL |
|
[ 李晗, 吴福忠, 杨万勤, 徐李亚, 倪祥银, 何洁, 常晨晖 ( 2015). 不同厚度雪被对高山森林6种凋落物分解过程中酸溶性和酸不溶性组分的影响. 生态学报, 35, 4687-4698.]
DOI URL |
|
| [16] |
Liu Y, Wang GL, Liu GB, Qu QL, Yuan ZC ( 2010). Difference and inherent linkage of root characteristics in different root classification of Pinus tabulaeformis seedlings. Chinese Journal of Plant Ecology, 34, 1386-1393.
DOI URL |
|
[ 刘莹, 王国梁, 刘国彬, 曲秋玲, 袁子成 ( 2010). 不同分类系统下油松幼苗根系特征的差异与联系. 植物生态学报, 34, 1386-1393.]
DOI URL |
|
| [17] |
Magill AH, John DA ( 1998). Long-term effects of experimental nitrogen additions on foliar litter decay and humus for mation in forest ecosystems. Plant and Soil, 203, 301-311.
DOI URL |
| [18] |
McCormack ML, Dickie IA, Eissenstat DM, Fahey TJ, Fernandez CW, Guo D, Helmisaari HS, Hobbie EA, Iversen CM, Jackson RB, Lepp?lammi-Kujansuu J, Norby RJ, Phillips RP, Pregitzer KS, Pritchard SG, Rewald B, Zadworny M ( 2015). Redefining fine roots improves understanding of belowground contributions to terrestrial biosphere processes. New Phytologist, 207, 505-518.
DOI URL PMID |
| [19] |
Pregitzer KS, de Forest JL, Burton AJ, Allen MF, Ruess RW, Hendrick RL ( 2002). Fine root architecture of nine North American trees. Ecological Monographs, 72, 293-309.
DOI URL |
| [20] |
Seifter S, Dayton S, Novic B ( 1950). The estimation of glycogen with the anthrone reagent. Archives of Biochemistry and Biophysics, 25, 191-200.
DOI URL PMID |
| [21] |
Silver WL, Miya RK ( 2001). Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia, 129, 407-419.
DOI URL PMID |
| [22] |
Song S, Gu JC, Quan XK, Guo DL, Wang ZQ ( 2008). Fine-root decomposition of Fraxinus mandschurica and Larix gmelinii plantations. Journal of Plant Ecology (Chinese Version), 32, 1227-1237.
DOI URL |
|
[ 宋森, 谷加存, 全先奎, 郭大立, 王政权 ( 2008). 水曲柳和兴安落叶松人工林细根分解研究. 植物生态学报, 32, 1227-1237.]
DOI URL |
|
| [23] |
Sun T, Dong L, Mao Z ( 2015). Simulated atmospheric nitrogen deposition alters decomposition of ephemeral roots. Ecosystems, 18, 1240-1252.
DOI URL |
| [24] | Sun T, Dong LL, Zhang LL, Wu ZJ, Wang QK, Li YY, Zhang HG, Wang ZW ( 2016). Early stage fine-root decomposition and its relationship with root order and soil depth in a Larix gmelinii plantation. Forests, 7, 234. DOI: 10.3390/?f7100234. |
| [25] |
Sun T, Mao ZZ, Dong LL, Hou LL, Song Y, Wang XW ( 2013a ). Further evidence for slow decomposition of very fine roots using two methods: Litterbags and intact cores. Plant and Soil, 366, 633-646.
DOI URL |
| [26] |
Sun T, Mao ZZ, Han YY ( 2013b ). Slow decomposition of very fine roots and some factors controlling the process: A 4-year experiment in four temperate tree species. Plant and Soil, 372, 445-458.
DOI URL |
| [27] |
Vogt KA, Grier CC, Vogt DJ ( 1986). Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests. Advances in Ecological Research, 15, 303-377.
DOI URL |
| [28] |
Wardle DA, Bardgett RD, Klironomos JN, Set?l? H, van der Putten WH, Wall WH ( 2004). Ecological linkages between aboveground and belowground biota. Science, 304, 1629-1633.
DOI URL |
| [29] |
Wen DZ, Wei P, Zhang YC, Kong GH ( 1998). Dry mass loss and chemical changes of the decomposed fine roots in three China south subtropical forests at Dinghushan. Chinese Journal of Ecology, 17, 1-6.
DOI URL |
|
[ 温达志, 魏平, 张佑昌, 孔国辉 ( 1998). 鼎湖山南亚热带森林细根分解干物质损失和元素动态. 生态学杂志, 17, 1-6.]
DOI URL |
|
| [30] | Xia M, Guo D, Pregitzer KS ( 2010). Ephemeral root modules in Fraxinus mandschurica. New Phytologist, 188, 1065-1074. |
| [31] |
Xia M, Talhelm AF, Pregitzer KS ( 2015). Fine roots are the dominant source of recalcitrant plant litter in sugar maple-?dominated northern hardwood forests. New Phytologist, 208, 715-726.
DOI URL PMID |
| [32] | Xiong Y, Fan P, Fu S, Zeng H, Guo D ( 2012). Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees. Plant and Soil, 363, 19-31. |
| [33] | Zhang XJ, Wu C, Mei L, Han YZ, Wang ZQ ( 2006). Root decomposition and nutrient release of Fraxinus manshurica and Larix gmelinii plantations. Chinese Journal of Applied Ecology, 17, 1370-1376. |
| [ 张秀娟, 吴楚, 梅莉, 韩有志, 王政权 ( 2006). 水曲柳和落叶松人工林细根分解与养分释放. 应用生态学报, 17, 1370-1376.] |
| [1] | 俞庆水 倪晓凤 吉成均 朱江玲 唐志尧 方精云. 10年氮磷添加对海南尖峰岭两种热带雨林优势植物叶片非结构性碳水化合物的影响[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 舒韦维, 杨坤, 马俊旭, 闵惠琳, 陈琳, 刘士玲, 黄日逸, 明安刚, 明财道, 田祖为. 氮添加对红锥不同序级细根形态和化学性状的影响[J]. 植物生态学报, 2024, 48(1): 103-112. |
| [3] | 苏炜, 陈平, 吴婷, 刘岳, 宋雨婷, 刘旭军, 刘菊秀. 氮添加与干季延长对降香黄檀幼苗非结构性碳水化合物、养分与生物量的影响[J]. 植物生态学报, 2023, 47(8): 1094-1104. |
| [4] | 余海霞, 曲鲁平, 汤行昊, 刘南, 张子雷, 王浩, 王艺璇, 邵长亮, 董刚, 胡亚林. 闽楠和木荷非结构性碳水化合物对不同模式热浪的差异性响应[J]. 植物生态学报, 2023, 47(2): 249-261. |
| [5] | 陈图强, 徐贵青, 刘深思, 李彦. 干旱胁迫下梭梭水力性状调整与非结构性碳水化合物动态[J]. 植物生态学报, 2023, 47(10): 1407-1421. |
| [6] | 李变变, 张凤华, 赵亚光, 孙秉楠. 不同刈割程度对油莎豆非结构性碳水化合物代谢及生物量的影响[J]. 植物生态学报, 2023, 47(1): 101-113. |
| [7] | 伍敏, 田雨, 樊大勇, 张祥雪. 干旱胁迫下毛白杨和元宝槭的水力学调控[J]. 植物生态学报, 2022, 46(9): 1086-1097. |
| [8] | 董涵君, 王兴昌, 苑丹阳, 柳荻, 刘玉龙, 桑英, 王晓春. 温带不同材性树种树干非结构性碳水化合物的径向分配差异[J]. 植物生态学报, 2022, 46(6): 722-734. |
| [9] | 李思源, 张照鑫, 饶良懿. 桑苗非结构性碳水化合物和生长激素对水淹胁迫的响应[J]. 植物生态学报, 2022, 46(3): 311-320. |
| [10] | 秦慧君, 焦亮, 周怡, 薛儒鸿, 柒常亮, 杜达石. 祁连山优势树木碳水化合物资源分配的海拔和树种效应[J]. 植物生态学报, 2022, 46(2): 208-219. |
| [11] | 林夏珍, 刘林, 董婷婷, 方琦博, 郭庆学. 非结构性碳水化合物与氮分配对美洲黑杨和青杨耐盐能力的影响[J]. 植物生态学报, 2021, 45(9): 961-971. |
| [12] | 吴秋霞, 吴福忠, 胡仪, 康自佳, 张耀艺, 杨静, 岳楷, 倪祥银, 杨玉盛. 亚热带同质园11个树种新老叶非结构性碳水化合物含量比较[J]. 植物生态学报, 2021, 45(7): 771-779. |
| [13] | 宋琳, 雒文涛, 马望, 何鹏, 梁潇洒, 王正文. 极端干旱对草甸草原优势植物非结构性碳水化合物的影响[J]. 植物生态学报, 2020, 44(6): 669-676. |
| [14] | 章异平, 海旭莹, 徐军亮, 吴文霞, 曹鹏鹤, 安文静. 秦岭东段栓皮栎枝条非结构性碳水化合物含量的季节动态[J]. 植物生态学报, 2019, 43(6): 521-531. |
| [15] | 周慧敏, 李品, 冯兆忠, 张殷波. 地表臭氧浓度升高与干旱交互作用对杨树非结构性碳水化合物积累和叶根分配的短期影响[J]. 植物生态学报, 2019, 43(4): 296-304. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19