
植物生态学报 ›› 2015, Vol. 39 ›› Issue (10): 980-989.DOI: 10.17521/cjpe.2015.0095
王立龙1, 王亮2, 张丽芳1, 刘玉洋1, 徐世健1,*( )
)
出版日期:2015-10-20
发布日期:2015-10-24
通讯作者:
徐世健
Li-Long WANG1, Liang WANG2, Li-Fang ZHANG1, Yu-Yang LIU1, Shi-Jian XU1,*()
Online:2015-10-20
Published:2015-10-24
Contact:
Shi-Jian XU
摘要:
裸果木(Gymnocarpos przewalskii)为古地中海旱生植物区系孑遗种, 是砾质荒漠的主要建群种之一。通过对安西极旱荒漠国家级自然保护区内3种生境(水冲滩地、山间冲沟、平缓戈壁)条件下裸果木种群的样地调查, 编制不同生境裸果木种群的静态生命表, 绘制其存活曲线和生存分析函数的曲线, 并采用时间序列模型对种群的数量动态进行预测, 以揭示不同生境下裸果木的种群结构和动态特征。结果表明: 3种生境下裸果木种群均属增长型, 数量变化动态指数(V’ pi) > 0, 但对外界干扰比较敏感, 存活曲线趋于Deevey-II型, 表明种群各龄级的死亡率基本接近; 不同生境裸果木种群大小依次为水冲滩地>山间冲沟>平缓戈壁, 结合对种群年龄结构的分析, 进一步表明水分条件较好的水冲滩地和山间冲沟生境更加适宜裸果木生存; 生存分析和时间序列预测表明幼龄个体的缺乏是未来裸果木种群发生衰退的主要原因, 且平缓戈壁生境下的裸果木种群将先于另外两种生境下的种群发生更加快速的衰退。
王立龙, 王亮, 张丽芳, 刘玉洋, 徐世健. 不同生境下濒危植物裸果木种群结构及动态特征. 植物生态学报, 2015, 39(10): 980-989. DOI: 10.17521/cjpe.2015.0095
Li-Long WANG, Liang WANG, Li-Fang ZHANG, Yu-Yang LIU, Shi-Jian XU. Structure and dynamic characteristics of Gymnocarpos przewalskii in different habitats. Chinese Journal of Plant Ecology, 2015, 39(10): 980-989. DOI: 10.17521/cjpe.2015.0095
| 生境 Habitat | 物种名称 Species name | 平均密度 Mean density (plants·plot-1) | 平均高度 Mean height (m) | 平均冠幅 Mean crown (m) | 平均盖度 Mean canopy (%) | 重要值 Important value | 群落组成 Community composition |
|---|---|---|---|---|---|---|---|
| 水冲滩地 Flooding land (20) | 1. 膜果麻黄 Ephedra przewalskii | 11.02 | 0.29 | 1.06 | 3.20 | 0.13 | 膜果麻黄 E. przewalskii + 裸果木 G. przewalskii + 合头草 S. regelii |
| 2. 裸果木 Gymnocarpos przewalskii | 10.60 | 0.34 | 0.74 | 3.10 | 0.12 | ||
| 3. 合头草 Sympegma regelii | 7.31 | 0.42 | 0.78 | 2.23 | 0.10 | ||
| 4. 红砂 Reaumuria songarica | 4.17 | 0.38 | 0.80 | 1.26 | 0.07 | ||
| 5. 细枝岩黄芪 Hedysarum scoparium | 1.61 | 0.62 | 1.18 | 0.66 | 0.06 | ||
| 山间冲沟 Gully (18) | 1. 合头草 Sympegma regelii | 13.80 | 0.43 | 0.80 | 3.00 | 0.19 | 合头草 S. regelii + 泡泡刺 N. sphaerocarpa + 裸果木 G. przewalskii |
| 2. 泡泡刺 Nitraria sphaerocarpa | 4.00 | 0.29 | 1.35 | 3.20 | 0.13 | ||
| 3. 裸果木 Gymnocarpos przewalskii | 4.10 | 0.36 | 0.85 | 3.00 | 0.12 | ||
| 4. 红砂 Reaumuria songarica | 2.80 | 0.27 | 0.68 | 0.80 | 0.06 | ||
| 5. 沙拐枣 Calligonum mongolicum | 0.41 | 0.56 | 1.17 | 0.42 | 0.05 | ||
| 平缓戈壁 Flat desert (18) | 1. 红砂 Reaumuria songarica | 11.80 | 0.26 | 0.75 | 2.20 | 0.14 | 红砂 R. songarica + 泡泡刺 N. sphaerocarpa + 合头草 S. regelii |
| 2. 泡泡刺 Nitraria sphaerocarpa | 7.02 | 0.27 | 0.91 | 1.81 | 0.11 | ||
| 3. 合头草 Sympegma regelii | 6.68 | 0.39 | 0.60 | 1.80 | 0.10 | ||
| 4. 膜果麻黄 Ephedra przewalskii | 2.91 | 0.37 | 0.95 | 1.92 | 0.09 | ||
| 5. 裸果木 Gymnocarpos przewalskii | 2.64 | 0.42 | 0.81 | 1.85 | 0.09 |
表1 不同生境下植物群落组成及其数量特征
Table 1 Community composition and quantity characteristics in different habitats
| 生境 Habitat | 物种名称 Species name | 平均密度 Mean density (plants·plot-1) | 平均高度 Mean height (m) | 平均冠幅 Mean crown (m) | 平均盖度 Mean canopy (%) | 重要值 Important value | 群落组成 Community composition |
|---|---|---|---|---|---|---|---|
| 水冲滩地 Flooding land (20) | 1. 膜果麻黄 Ephedra przewalskii | 11.02 | 0.29 | 1.06 | 3.20 | 0.13 | 膜果麻黄 E. przewalskii + 裸果木 G. przewalskii + 合头草 S. regelii |
| 2. 裸果木 Gymnocarpos przewalskii | 10.60 | 0.34 | 0.74 | 3.10 | 0.12 | ||
| 3. 合头草 Sympegma regelii | 7.31 | 0.42 | 0.78 | 2.23 | 0.10 | ||
| 4. 红砂 Reaumuria songarica | 4.17 | 0.38 | 0.80 | 1.26 | 0.07 | ||
| 5. 细枝岩黄芪 Hedysarum scoparium | 1.61 | 0.62 | 1.18 | 0.66 | 0.06 | ||
| 山间冲沟 Gully (18) | 1. 合头草 Sympegma regelii | 13.80 | 0.43 | 0.80 | 3.00 | 0.19 | 合头草 S. regelii + 泡泡刺 N. sphaerocarpa + 裸果木 G. przewalskii |
| 2. 泡泡刺 Nitraria sphaerocarpa | 4.00 | 0.29 | 1.35 | 3.20 | 0.13 | ||
| 3. 裸果木 Gymnocarpos przewalskii | 4.10 | 0.36 | 0.85 | 3.00 | 0.12 | ||
| 4. 红砂 Reaumuria songarica | 2.80 | 0.27 | 0.68 | 0.80 | 0.06 | ||
| 5. 沙拐枣 Calligonum mongolicum | 0.41 | 0.56 | 1.17 | 0.42 | 0.05 | ||
| 平缓戈壁 Flat desert (18) | 1. 红砂 Reaumuria songarica | 11.80 | 0.26 | 0.75 | 2.20 | 0.14 | 红砂 R. songarica + 泡泡刺 N. sphaerocarpa + 合头草 S. regelii |
| 2. 泡泡刺 Nitraria sphaerocarpa | 7.02 | 0.27 | 0.91 | 1.81 | 0.11 | ||
| 3. 合头草 Sympegma regelii | 6.68 | 0.39 | 0.60 | 1.80 | 0.10 | ||
| 4. 膜果麻黄 Ephedra przewalskii | 2.91 | 0.37 | 0.95 | 1.92 | 0.09 | ||
| 5. 裸果木 Gymnocarpos przewalskii | 2.64 | 0.42 | 0.81 | 1.85 | 0.09 |
| 种群动态指数级 Dynamic index | 动态指数 Dynamic index value (%) | ||
|---|---|---|---|
| 水冲滩地 Flooding land | 山间冲沟 Gully | 平缓戈壁 Flat desert | |
| V1 | -26.67 | -40.74 | -60.00 |
| V2 | -10.45 | 40.74 | -77.27 |
| V3 | -5.63 | -23.81 | -37.14 |
| V4 | 11.27 | -30.00 | 20.00 |
| V5 | 31.75 | 46.67 | 0.00 |
| V6 | 60.47 | 31.25 | 71.43 |
| V7 | 35.29 | 36.36 | 75.00 |
| V8 | 72.73 | 85.71 | 0.00 |
| V9 | 66.67 | -66.67 | 50.00 |
| Vpi | 12.72 | 15.66 | 15.73 |
| V' pi | 1.27 | 1.57 | 1.57 |
表2 不同生境下裸果木种群龄级结构动态变化指数
Table 2 Dynamic indices of Gymnocarpos przewalskii population and size structure in different habitats
| 种群动态指数级 Dynamic index | 动态指数 Dynamic index value (%) | ||
|---|---|---|---|
| 水冲滩地 Flooding land | 山间冲沟 Gully | 平缓戈壁 Flat desert | |
| V1 | -26.67 | -40.74 | -60.00 |
| V2 | -10.45 | 40.74 | -77.27 |
| V3 | -5.63 | -23.81 | -37.14 |
| V4 | 11.27 | -30.00 | 20.00 |
| V5 | 31.75 | 46.67 | 0.00 |
| V6 | 60.47 | 31.25 | 71.43 |
| V7 | 35.29 | 36.36 | 75.00 |
| V8 | 72.73 | 85.71 | 0.00 |
| V9 | 66.67 | -66.67 | 50.00 |
| Vpi | 12.72 | 15.66 | 15.73 |
| V' pi | 1.27 | 1.57 | 1.57 |
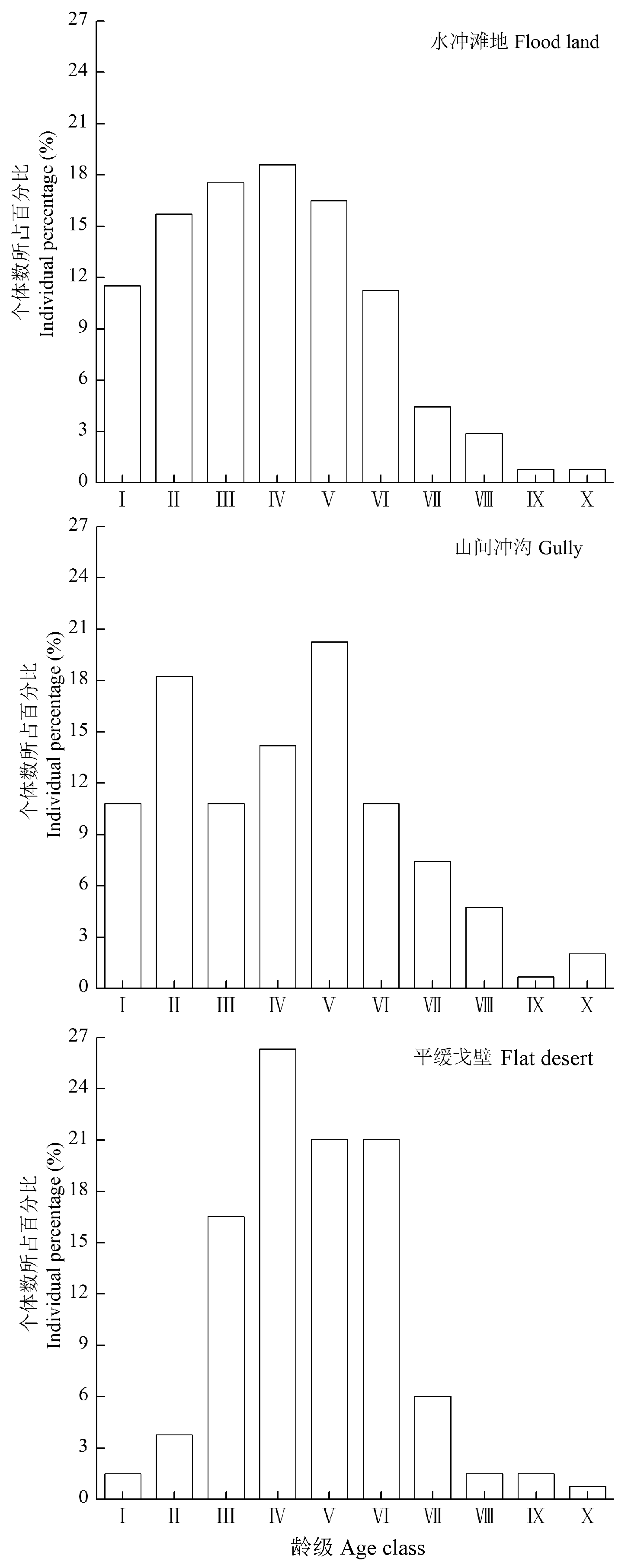
图1 不同生境下裸果木种群的龄级结构。采用高度级(H)结构代替年龄结构, 以10 cm为间隔共划分10个龄级, I (H < 10 cm)、II (10 cm ≤ H < 20 cm)……X (H ≥ 100 cm)。
Fig. 1 The age structure of Gymnocarpos przewalskii population in different habitats. The age structure was replaced by height-class structure (H), totally 10 age classes were divided in intervals of 10 cm, I (H < 10 cm), II (10 cm ≤ H < 20 cm) …, X (H ≥ 100 cm).
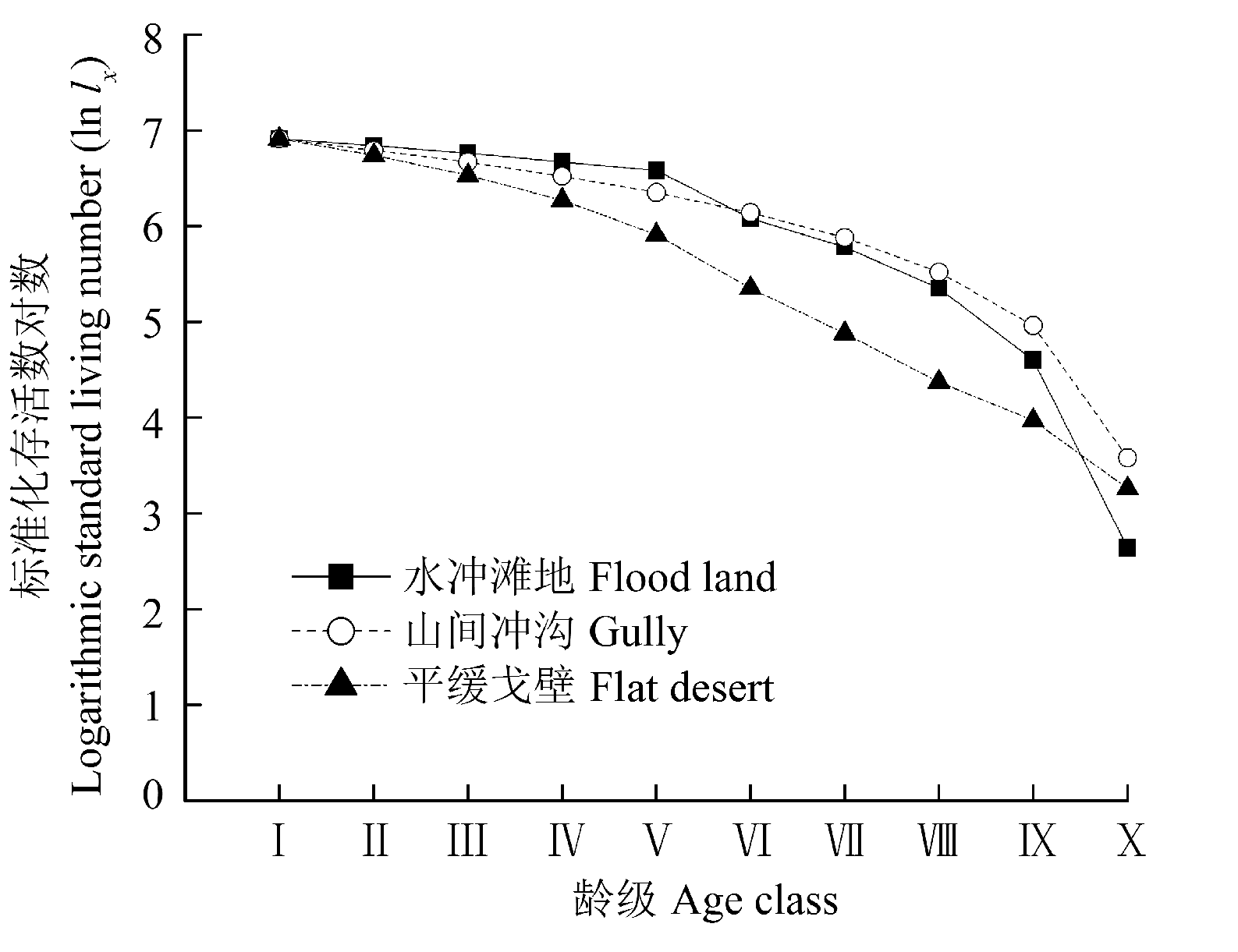
图2 不同生境下裸果木种群的存活曲线。龄级同图1。
Fig. 2 Survival curve of Gymnocarpos przewalskii population in different habitats. Age class see Fig. 1.
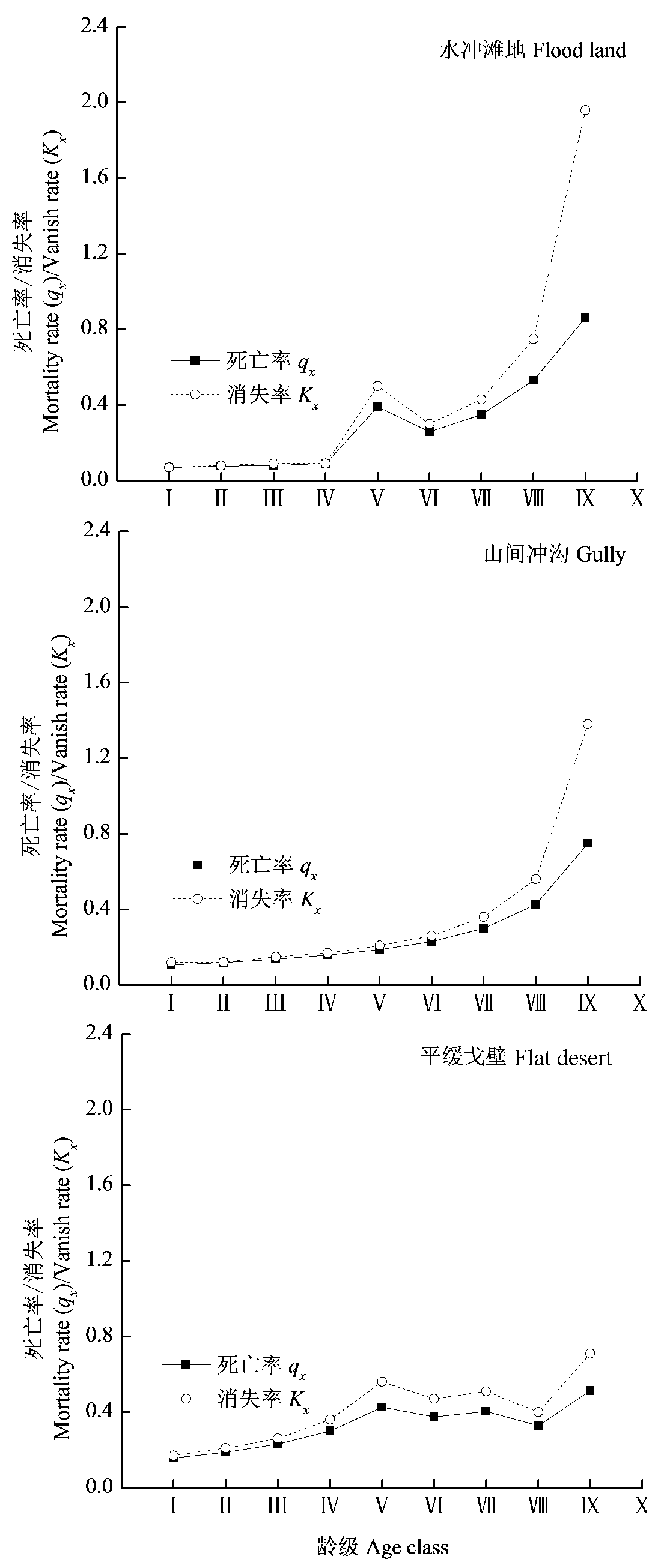
图3 不同生境下裸果木种群的死亡率和消失率。龄级同图1。
Fig. 3 Mortality rate and disappearance rate of Gymnocarpos przewalskii population in different habitats. Age class see Fig. 1.
| 生境 Habitat | 龄级 Age class | 高度 Height (cm) | Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 水冲滩地 Flooding land | I | H < 10 | 44 | 71 | 1 000 | 70 | 0.07 | 965 | 4 876 | 4.88 | 6.91 | 0.07 |
| II | 10 ≤ H < 20 | 60 | 66 | 930 | 71 | 0.08 | 895 | 3 911 | 4.21 | 6.84 | 0.08 | |
| III | 20 ≤ H < 30 | 67 | 61 | 859 | 70 | 0.08 | 824 | 3 016 | 3.51 | 6.76 | 0.09 | |
| IV | 30 ≤ H < 40 | 71 | 56 | 789 | 71 | 0.09 | 753 | 2 192 | 2.78 | 6.67 | 0.09 | |
| V | 40 ≤ H < 50 | 63 | 51 | 718 | 281 | 0.39 | 578 | 1 439 | 2.00 | 6.58 | 0.50 | |
| VI | 50 ≤ H < 60 | 43 | 31 | 437 | 113 | 0.26 | 381 | 861 | 1.97 | 6.08 | 0.30 | |
| VII | 60 ≤ H < 70 | 17 | 23 | 324 | 113 | 0.35 | 268 | 480 | 1.48 | 5.78 | 0.43 | |
| VIII | 80 ≤ H < 90 | 11 | 15 | 211 | 112 | 0.53 | 155 | 212 | 1.00 | 5.35 | 0.75 | |
| IX | 90 ≤ H < 100 | 3 | 7 | 99 | 85 | 0.86 | 57 | 57 | 0.58 | 4.60 | 1.96 | |
| X | H ≥ 100 | 3 | 1 | 14 | - | - | - | - | - | 2.64 | - | |
| 山间冲沟 Gully | I | H < 10 | 16 | 28 | 1 000 | 107 | 0.11 | 946 | 4 661 | 4.66 | 6.91 | 0.12 |
| II | 10 ≤ H < 20 | 27 | 25 | 893 | 107 | 0.12 | 839 | 3 714 | 4.16 | 6.79 | 0.12 | |
| III | 20 ≤ H < 30 | 16 | 22 | 786 | 107 | 0.14 | 732 | 2 875 | 3.66 | 6.67 | 0.15 | |
| IV | 30 ≤ H < 40 | 21 | 19 | 679 | 108 | 0.16 | 625 | 2 143 | 3.16 | 6.52 | 0.17 | |
| V | 40 ≤ H < 50 | 30 | 16 | 571 | 107 | 0.19 | 518 | 1 518 | 2.66 | 6.35 | 0.21 | |
| VI | 50 ≤ H < 60 | 16 | 13 | 464 | 107 | 0.23 | 411 | 1 000 | 2.15 | 6.14 | 0.26 | |
| VII | 60 ≤ H < 70 | 11 | 10 | 357 | 107 | 0.30 | 304 | 589 | 1.65 | 5.88 | 0.36 | |
| VIII | 80 ≤ H < 90 | 7 | 7 | 250 | 107 | 0.43 | 196 | 286 | 1.14 | 5.52 | 0.56 | |
| IX | 90 ≤ H < 100 | 0 | 4 | 143 | 107 | 0.75 | 89 | 89 | 0.63 | 4.96 | 1.38 | |
| X | H ≥ 100 | 3 | 1 | 36 | - | - | - | - | - | 3.58 | - | |
| 平缓戈壁 Flat desert | I | H < 10 | 2 | 38 | 1 000 | 158 | 0.16 | 921 | 3 408 | 3.41 | 6.91 | 0.17 |
| II | 10 ≤ H < 20 | 5 | 32 | 842 | 158 | 0.19 | 763 | 2 487 | 2.95 | 6.74 | 0.21 | |
| III | 20 ≤ H < 30 | 22 | 26 | 684 | 158 | 0.23 | 605 | 1 724 | 2.52 | 6.53 | 0.26 | |
| IV | 30 ≤ H < 40 | 35 | 20 | 526 | 158 | 0.30 | 447 | 1 118 | 2.13 | 6.27 | 0.36 | |
| V | 40 ≤ H < 50 | 28 | 14 | 368 | 157 | 0.43 | 289 | 671 | 1.82 | 5.91 | 0.56 | |
| VI | 50 ≤ H < 60 | 28 | 8 | 211 | 79 | 0.38 | 171 | 382 | 1.81 | 5.35 | 0.47 | |
| VII | 60 ≤ H < 70 | 8 | 5 | 132 | 53 | 0.40 | 105 | 211 | 1.60 | 4.88 | 0.51 | |
| VIII | 80 ≤ H < 90 | 2 | 3 | 79 | 26 | 0.33 | 66 | 105 | 1.33 | 4.37 | 0.40 | |
| IX | 90 ≤ H < 100 | 2 | 2 | 53 | 27 | 0.51 | 39 | 39 | 0.75 | 3.97 | 0.71 | |
| X | H ≥ 100 | 1 | 1 | 26 | - | - | - | - | - | 3.26 | - |
表3 不同生境下裸果木种群静态生命表
Table 3 Static life table of Gymnocarpos przewalskii population in different habitats
| 生境 Habitat | 龄级 Age class | 高度 Height (cm) | Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 水冲滩地 Flooding land | I | H < 10 | 44 | 71 | 1 000 | 70 | 0.07 | 965 | 4 876 | 4.88 | 6.91 | 0.07 |
| II | 10 ≤ H < 20 | 60 | 66 | 930 | 71 | 0.08 | 895 | 3 911 | 4.21 | 6.84 | 0.08 | |
| III | 20 ≤ H < 30 | 67 | 61 | 859 | 70 | 0.08 | 824 | 3 016 | 3.51 | 6.76 | 0.09 | |
| IV | 30 ≤ H < 40 | 71 | 56 | 789 | 71 | 0.09 | 753 | 2 192 | 2.78 | 6.67 | 0.09 | |
| V | 40 ≤ H < 50 | 63 | 51 | 718 | 281 | 0.39 | 578 | 1 439 | 2.00 | 6.58 | 0.50 | |
| VI | 50 ≤ H < 60 | 43 | 31 | 437 | 113 | 0.26 | 381 | 861 | 1.97 | 6.08 | 0.30 | |
| VII | 60 ≤ H < 70 | 17 | 23 | 324 | 113 | 0.35 | 268 | 480 | 1.48 | 5.78 | 0.43 | |
| VIII | 80 ≤ H < 90 | 11 | 15 | 211 | 112 | 0.53 | 155 | 212 | 1.00 | 5.35 | 0.75 | |
| IX | 90 ≤ H < 100 | 3 | 7 | 99 | 85 | 0.86 | 57 | 57 | 0.58 | 4.60 | 1.96 | |
| X | H ≥ 100 | 3 | 1 | 14 | - | - | - | - | - | 2.64 | - | |
| 山间冲沟 Gully | I | H < 10 | 16 | 28 | 1 000 | 107 | 0.11 | 946 | 4 661 | 4.66 | 6.91 | 0.12 |
| II | 10 ≤ H < 20 | 27 | 25 | 893 | 107 | 0.12 | 839 | 3 714 | 4.16 | 6.79 | 0.12 | |
| III | 20 ≤ H < 30 | 16 | 22 | 786 | 107 | 0.14 | 732 | 2 875 | 3.66 | 6.67 | 0.15 | |
| IV | 30 ≤ H < 40 | 21 | 19 | 679 | 108 | 0.16 | 625 | 2 143 | 3.16 | 6.52 | 0.17 | |
| V | 40 ≤ H < 50 | 30 | 16 | 571 | 107 | 0.19 | 518 | 1 518 | 2.66 | 6.35 | 0.21 | |
| VI | 50 ≤ H < 60 | 16 | 13 | 464 | 107 | 0.23 | 411 | 1 000 | 2.15 | 6.14 | 0.26 | |
| VII | 60 ≤ H < 70 | 11 | 10 | 357 | 107 | 0.30 | 304 | 589 | 1.65 | 5.88 | 0.36 | |
| VIII | 80 ≤ H < 90 | 7 | 7 | 250 | 107 | 0.43 | 196 | 286 | 1.14 | 5.52 | 0.56 | |
| IX | 90 ≤ H < 100 | 0 | 4 | 143 | 107 | 0.75 | 89 | 89 | 0.63 | 4.96 | 1.38 | |
| X | H ≥ 100 | 3 | 1 | 36 | - | - | - | - | - | 3.58 | - | |
| 平缓戈壁 Flat desert | I | H < 10 | 2 | 38 | 1 000 | 158 | 0.16 | 921 | 3 408 | 3.41 | 6.91 | 0.17 |
| II | 10 ≤ H < 20 | 5 | 32 | 842 | 158 | 0.19 | 763 | 2 487 | 2.95 | 6.74 | 0.21 | |
| III | 20 ≤ H < 30 | 22 | 26 | 684 | 158 | 0.23 | 605 | 1 724 | 2.52 | 6.53 | 0.26 | |
| IV | 30 ≤ H < 40 | 35 | 20 | 526 | 158 | 0.30 | 447 | 1 118 | 2.13 | 6.27 | 0.36 | |
| V | 40 ≤ H < 50 | 28 | 14 | 368 | 157 | 0.43 | 289 | 671 | 1.82 | 5.91 | 0.56 | |
| VI | 50 ≤ H < 60 | 28 | 8 | 211 | 79 | 0.38 | 171 | 382 | 1.81 | 5.35 | 0.47 | |
| VII | 60 ≤ H < 70 | 8 | 5 | 132 | 53 | 0.40 | 105 | 211 | 1.60 | 4.88 | 0.51 | |
| VIII | 80 ≤ H < 90 | 2 | 3 | 79 | 26 | 0.33 | 66 | 105 | 1.33 | 4.37 | 0.40 | |
| IX | 90 ≤ H < 100 | 2 | 2 | 53 | 27 | 0.51 | 39 | 39 | 0.75 | 3.97 | 0.71 | |
| X | H ≥ 100 | 1 | 1 | 26 | - | - | - | - | - | 3.26 | - |
| 生境 Habitat | 方程 Equation | R2 | F | p |
|---|---|---|---|---|
| 水冲滩地 Flooding land | Nx = 8.718e-0.079x | 0.660 | 15.52 | 0.004 |
| Nx = 8.409x-0.266 | 0.432 | 6.076 | 0.039 | |
| 山间冲沟 Gully | Nx = 7.986e-0.057x | 0.745 | 23.42 | 0.001 |
| Nx = 7.858x-0.197 | 0.521 | 8.711 | 0.018 | |
| 平缓戈壁 Flat desert | Nx = 8.223e-0.081x | 0.935 | 115.6 | 0.000 |
| Nx = 8.216x-0.294 | 0.724 | 21.02 | 0.002 |
表4 不同生境下裸果木种群存活曲线的检验模型
Table 4 Test models of survival curves of Gymnocarpos przewalskii in different habitats
| 生境 Habitat | 方程 Equation | R2 | F | p |
|---|---|---|---|---|
| 水冲滩地 Flooding land | Nx = 8.718e-0.079x | 0.660 | 15.52 | 0.004 |
| Nx = 8.409x-0.266 | 0.432 | 6.076 | 0.039 | |
| 山间冲沟 Gully | Nx = 7.986e-0.057x | 0.745 | 23.42 | 0.001 |
| Nx = 7.858x-0.197 | 0.521 | 8.711 | 0.018 | |
| 平缓戈壁 Flat desert | Nx = 8.223e-0.081x | 0.935 | 115.6 | 0.000 |
| Nx = 8.216x-0.294 | 0.724 | 21.02 | 0.002 |
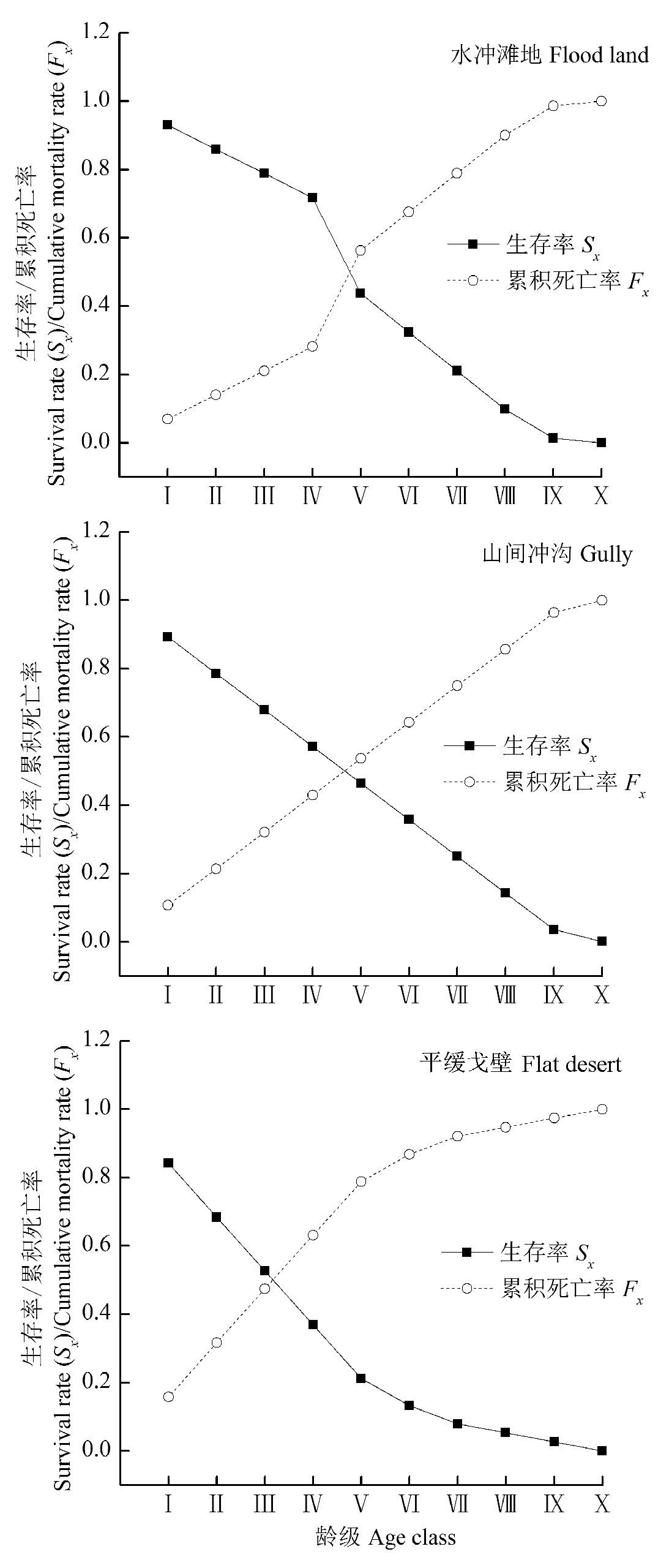
图4 不同生境下裸果木种群生存率和累计死亡率。龄级同图1。
Fig. 4 Survival rate and cumulative mortality rate of Gymnocarpos przewalskii population in different habitats. Age class see Fig. 1.
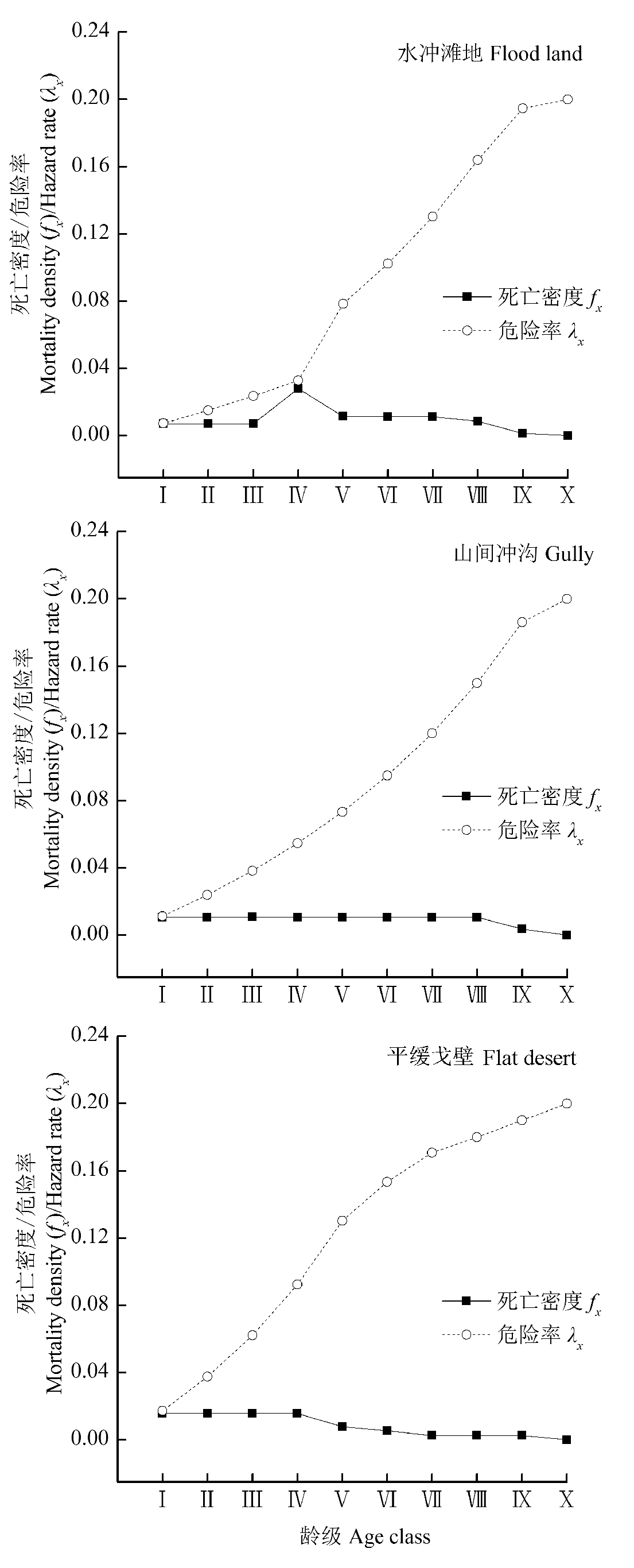
图5 不同生境下裸果木种群死亡密度和危险率。龄级同图1。
Fig. 5 Mortality density and hazard rate of Gymnocarpos przewalskii population in different habitats. Age class see Fig. 1.
| 龄级 Age class | 水冲滩地 Flood land | 山间冲沟 Gully | 平缓戈壁 Flat desert | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 原始数据 Primary data | M2(1) | M4(1) | M6(1) | M8(1) | 原始数据 Primary data | M2(1) | M4(1) | M6(1) | M8(1) | 原始数据 Primary data | M2(1) | M4(1) | M6(1) | M8(1) | |||
| I | 44 | 16 | 2 | ||||||||||||||
| II | 60 | 52 | 27 | 22 | 5 | 4 | |||||||||||
| III | 67 | 64 | 16 | 22 | 22 | 14 | |||||||||||
| IV | 71 | 69 | 61 | 21 | 19 | 20 | 35 | 29 | 16 | ||||||||
| V | 63 | 67 | 65 | 30 | 26 | 24 | 28 | 32 | 23 | ||||||||
| VI | 43 | 53 | 61 | 58 | 16 | 23 | 21 | 21 | 28 | 28 | 28 | 20 | |||||
| VII | 17 | 30 | 49 | 54 | 11 | 14 | 20 | 20 | 8 | 18 | 25 | 21 | |||||
| VIII | 11 | 14 | 34 | 45 | 47 | 7 | 9 | 16 | 17 | 18 | 2 | 5 | 17 | 21 | 16 | ||
| IX | 3 | 7 | 19 | 52 | 42 | 1 | 4 | 9 | 14 | 16 | 2 | 2 | 10 | 17 | 16 | ||
| X | 3 | 3 | 9 | 23 | 35 | 3 | 2 | 6 | 11 | 13 | 1 | 2 | 3 | 12 | 16 | ||
表5 不同生境下裸果木种群动态变化的时间序列分析
Table 5 Time sequence analysis of age structure of Gymnocarpos przewalskii population under different habitats
| 龄级 Age class | 水冲滩地 Flood land | 山间冲沟 Gully | 平缓戈壁 Flat desert | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 原始数据 Primary data | M2(1) | M4(1) | M6(1) | M8(1) | 原始数据 Primary data | M2(1) | M4(1) | M6(1) | M8(1) | 原始数据 Primary data | M2(1) | M4(1) | M6(1) | M8(1) | |||
| I | 44 | 16 | 2 | ||||||||||||||
| II | 60 | 52 | 27 | 22 | 5 | 4 | |||||||||||
| III | 67 | 64 | 16 | 22 | 22 | 14 | |||||||||||
| IV | 71 | 69 | 61 | 21 | 19 | 20 | 35 | 29 | 16 | ||||||||
| V | 63 | 67 | 65 | 30 | 26 | 24 | 28 | 32 | 23 | ||||||||
| VI | 43 | 53 | 61 | 58 | 16 | 23 | 21 | 21 | 28 | 28 | 28 | 20 | |||||
| VII | 17 | 30 | 49 | 54 | 11 | 14 | 20 | 20 | 8 | 18 | 25 | 21 | |||||
| VIII | 11 | 14 | 34 | 45 | 47 | 7 | 9 | 16 | 17 | 18 | 2 | 5 | 17 | 21 | 16 | ||
| IX | 3 | 7 | 19 | 52 | 42 | 1 | 4 | 9 | 14 | 16 | 2 | 2 | 10 | 17 | 16 | ||
| X | 3 | 3 | 9 | 23 | 35 | 3 | 2 | 6 | 11 | 13 | 1 | 2 | 3 | 12 | 16 | ||
| [1] | Chai YQ, Cao ZZ, Chai ZS (2010). A study on species diversity of a Gymnocarpos przewalskii community in the Subei desert.Acta Prataculturae Sinica ,19(1), 21-27. (in Chinese with English abstract) |
| [柴永青, 曹致中, 蔡卓山 (2010). 肃北地区裸果木荒漠群落构成特征及物种多样性研究. 草业学报, 19(1), 21-27.] | |
| [2] | Chen XD (1999). A study on the method of quantitative analysis for plant population and community structural dynamics.Acta Ecologica Sinica, 18, 214-217. (in Chinese with English abstract) |
| [陈晓德 (1999). 植物种群与群落结构动态量化分析方法研究. 生态学报, 18, 214-217.] | |
| [3] | Deevey ES Jr (1947). Life tables for natural populations of animals.The Quarterly Review of Biology, 22, 283-314. |
| [4] | Gurevitch J, Scheiner SM, Fox GA (2002). The Ecology of Plants. Sinauer Associates, Sunderland, Massachusetts, USA. |
| [5] | Harcombe PA (1987). Tree life tables.Bioscience, 37, 557-568. |
| [6] | Hett JM, Loucks OL (1976). Age structure models of balsam fir and eastern hemlock.The Journal of Ecology, 64, 1029-1044. |
| [7] | Hong W, Wang XG, Wu CZ, He JD, Liao CZ, Cheng Y, Feng L (2004). Life table and spectral analysis of endangered plant Taxus chinensis var. mairei population.Chinese Journal of Applied Ecology ,15, 1109-1112. (in Chinese with English abstract) |
| [洪伟, 王新功, 吴承祯, 何东进, 廖成章, 程煜, 封磊 (2004). 濒危植物南方红豆杉种群生命表及谱分析. 应用生态学报, 15, 1109-1112.] | |
| [8] | Jiang H (1992). Population Ecology of Spruce. China Forestry Publishing House, Beijing. (in Chinese) |
| [江洪 (1992). 云杉种群生态学. 中国林业出版社, 北京.] | |
| [9] | Ma DH (2006). Study on water resources development and utilization of Shule River Basin.Water Resources and Hydropower Engineering, 37(4), 1-4. (in Chinese with English abstract) |
| [马德海 (2006). 疏勒河流域水资源开发利用研究. 水利水电技术, 37(4), 1-4.] | |
| [10] | Ma SM, Zhang ML, Sanderson SC (2012). Phylogeography of the rare Gymnocarpos przewalskii(Caryophyllaceae): Indications of multiple glacial refugia in north-western China.Australian Journal of Botany ,60, 20-31. |
| [11] | Ma SM, Zhang ML, Zhang HX, Meng HH, Chen X (2010). Predicting potential geographical distributions and patterns of the relic plant Gymnocarpos przewalskii using maximum entropy and genetic algorithm for rule-set prediction.Chinese Journal of Plant Ecology ,34, 1327-1335. (in Chinese with English abstract) |
| [马松梅, 张明理, 张宏祥, 孟宏虎, 陈曦 (2010). 利用最大熵模型和规则集遗传算法模型预测孑遗植物裸果木的潜在地理分布及格局. 植物生态学报, 34, 1327-1335.] | |
| [12] | Ma SM, Zhang ML. (2012). Phylogeography and conservation genetics of the relic Gymnocarpos przewalskii(Caryophyllaceae) restricted to northwestern China.Conservation Genetics ,13, 1531-1541. |
| [13] | Shen SK, Ma HY, Wang YH, Wang BY, Shen GZ (2008). The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang.Acta Ecologica Sinica ,28, 2404-2412. (in Chinese with English abstract) |
| [申仕康, 马海英, 王跃华, 王博轶, 申国柱 (2008). 濒危植物猪血木(Euryodendron excelsum H. T. Chang)自然种群结构及动态. 生态学报, 28, 2404-2412.] | |
| [14] | Wang W, Liu CR, Ma KP, Yu SL (1999). Population structure and dynamics of Quercus liaotungensis in two broad-leaved deciduous forests in Dongling Mountain, northern China.Acta Botanica Sinica ,41, 425-432. (in Chinese with English abstract) |
| [王巍, 刘灿然, 马克平, 于顺利 (1999). 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 41, 425-432.] | |
| [15] | Wang ZB, Zhou XJ (2010). Research progress on rare endangered plant Gymnocarpos przewalskii.Resource Development & Market ,26, 436-437, 446. (in Chinese with English abstract) |
| [汪之波, 周向军 (2010). 珍稀濒危植物裸果木研究进展. 资源开发与市场, 26, 436-437, 446.] | |
| [16] | Wu CZ, Hong W, Xie JS, Wu JL (2000). Life table analysis of Tsuga longibracteata population.Chinese Journal of Applied Ecology ,11, 333-336. (in Chinese with English abstract) |
| [吴承祯, 洪伟, 谢金寿, 吴继林 (2000). 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 11, 333-336.] | |
| [17] | Xiao YA, He P, Li XH, Deng HP (2004). Study on numeric dynamics of natural populations of the endangered species Disanthus cercidifolius var. longipes.Acta Phytoecologica Sinica ,28, 252-257. (in Chinese with English abstract) |
| [肖宜安, 何平, 李晓红, 邓洪平 (2004). 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 28, 252-257.] | |
| [18] | Xie TT, Su PX, Zhou ZJ, Zhang HN, Li SJ (2014). Structure and dynamic characteristics of Calligonum mongolicum population in the desert-oasis ecotone.Acta Ecologica Sinica ,34, 4272-4279. (in Chinese with English abstract) |
| [解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家 (2014). 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 34, 4272-4279.] | |
| [19] | Xie ZQ, Chen WL, Lu P, Hu D (1999). The demography and age structure of the endangered plant population of Cathaya argyrophylla.Acta Ecologica Sinica ,19, 523-528. (in Chinese with English abstract) |
| [谢宗强, 陈伟烈, 路鹏, 胡东 (1999). 濒危植物银杉的种群统计与年龄结构. 生态学报, 19, 523-528.] | |
| [20] | Yang FX, Wang SQ, Xu HG, Li BQ (1991). The theory of survival analysis and its application to life table.Acta Ecologica Sinica, 11, 153-158. (in Chinese with English abstract) |
| [杨凤翔, 王顺庆, 徐海根, 李邦庆 (1991). 生存分析理论及其在研究生命表中的应用. 生态学报, 11, 153-158.] | |
| [21] | Yang JM, Chen XL, Gong DJ (2013). Study on species diversity of rare endangered plant Gymnocarpos przewalskii communities.Journal of Anhui Agricultural Sciences ,41, 8508-8511. (in Chinese with English abstract) |
| [杨建美, 陈学林, 龚大洁 (2013). 珍稀濒危植物裸果木群落物种多样性研究. 安徽农业科学, 41, 8508-8511.] | |
| [22] | Yi XM, Zhang Y, Ji LZ (2013). Population structure of Phellodendron amurense in Changbai Mountains of northeast China.Chinese Journal of Ecology ,32, 2257-2262. |
| [易雪梅, 张悦, 姬兰柱 (2013). 长白山黄檗种群结构. 生态学杂志, 32, 2257-2262.] | |
| [23] | Zheng GZ, Yue LP, He JF, Wang JX, Zhang YL (2006). Grain-size characteristics of the sediments at Palaeoswamp in Anxi County in downstream of Shulehe River during Holocene and its paleoclimatic significance.Acta Sedimentologica Sinica, 24, 733-739. (in Chinese with English abstract) |
| [郑国璋, 岳乐平, 何军锋, 王建新, 张玉玲 (2006). 疏勒河下游安西古沼泽全新世沉积物粒度特征及其古气候环境意义. 沉积学报, 24, 733-739.] |
| [1] | 文佳 张新娜 王娟 赵秀海 张春雨. 性状调节幼苗存活率对邻体竞争和环境的响应 [J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 马常钦, 黄海龙, 彭政淋, 吴纯泽, 韦庆钰, 贾红涛, 卫星. 水曲柳雌雄株复叶类型及光合功能对不同生境的响应[J]. 植物生态学报, 2023, 47(9): 1287-1297. |
| [3] | 王燕玲, 招礼军, 朱栗琼, 莫若果, 林婷, 赵小雨. 广西天然红鳞蒲桃种群幼苗数量特征及动态分析[J]. 植物生态学报, 2023, 47(9): 1278-1286. |
| [4] | 冯珊珊, 黄春晖, 唐梦云, 蒋维昕, 白天道. 细叶云南松针叶形态和显微性状地理变异及其环境解释[J]. 植物生态学报, 2023, 47(8): 1116-1130. |
| [5] | 管岳, 王妍欣, 褚佳瑶, 冯琳骄, 宋晓萌, 周龙. 新疆野扁桃种群年龄结构及动态分析[J]. 植物生态学报, 2023, 47(7): 967-977. |
| [6] | 石荡, 郭传超, 蒋南林, 唐莹莹, 郑凤, 王瑾, 廖康, 刘立强. 新疆野杏天然更新幼株的个体特征及空间分布格局[J]. 植物生态学报, 2023, 47(4): 515-529. |
| [7] | 冯可, 刘冬梅, 张琦, 安菁, 何双辉. 旅游干扰对松山油松林土壤微生物多样性及群落结构的影响[J]. 植物生态学报, 2023, 47(4): 584-596. |
| [8] | 汪晶晶, 王嘉浩, 黄致云, Vanessa Chiamaka OKECHUKW, 胡蝶, 祁珊珊, 戴志聪, 杜道林. 不同氮水平下内生固氮菌对入侵植物南美蟛蜞菊生长策略的影响[J]. 植物生态学报, 2023, 47(2): 195-205. |
| [9] | 林春惠, 顾惠怡, 叶钦良, 张志坚, 钟智明, 易绮斐. 珍稀濒危植物大苞山茶种群结构与动态特征[J]. 植物生态学报, 2023, 47(12): 1684-1692. |
| [10] | 何春梅, 李雨姗, 尹秋龙, 贾仕宏, 郝占庆. 秦岭皇冠暖温性落叶阔叶林优势树种的径级结构和数量特征[J]. 植物生态学报, 2023, 47(12): 1658-1667. |
| [11] | 张金峰, 葛树森, 梁金花, 李俊清. 长白山阔叶红松林红松种群年龄结构与数量动态特征[J]. 植物生态学报, 2022, 46(6): 667-677. |
| [12] | 余秋伍, 杨菁, 沈国春. 浙江天童常绿阔叶林林冠结构与群落物种组成的关系[J]. 植物生态学报, 2022, 46(5): 529-538. |
| [13] | 孟庆静, 樊卫国. 刺梨的适钙类型及对高钙生境的适应性[J]. 植物生态学报, 2022, 46(12): 1562-1572. |
| [14] | 钟雨辰, 王斌, 方中平, 徐小忠, 于明坚. 片段化景观中壳斗科植物种子捕食和扩散模式[J]. 植物生态学报, 2021, 45(2): 154-162. |
| [15] | 韩大勇, 张维, 努尔买买提•依力亚斯, 杨允菲. 植物种群更新的补充限制[J]. 植物生态学报, 2021, 45(1): 1-12. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19