
植物生态学报 ›› 2005, Vol. 29 ›› Issue (6): 954-960.DOI: 10.17521/cjpe.2005.0124
姚红, 谭敦炎*( )
)
收稿日期:2005-01-20
接受日期:2005-04-15
出版日期:2005-01-20
发布日期:2005-09-30
通讯作者:
谭敦炎
作者简介:*E-mail:tandunyan@163.com基金资助:
YAO Hong, TAN Dun-Yan*()
Received:2005-01-20
Accepted:2005-04-15
Online:2005-01-20
Published:2005-09-30
Contact:
TAN Dun-Yan
摘要:
对准噶尔荒漠中胡卢巴属(Trigonella)4种短命植物网脉胡卢巴(T. cancellata)、单花胡卢巴(T. monantha)、直果胡卢巴(T. orthoceras)和弯果胡卢巴(T. arcuata)的物候特征及生物量分配进行了比较研究,结果表明:1)这4种植物的萌发对策多样,具有春、夏、秋萌现象,但以春、秋季萌发为主。其萌发时间在种间差异不明显,而与温度、降水等环境因子密切相关;2)生长发育快,生活周期短,一次结实,在干热夏季来临前完成生活周期,因此属于典型的避旱型植物;3) 秋、春萌株在早春返青或种子萌发后,经过短暂的营养生长后很快进入生殖生长,通过物候期(尤其是营养生长与生殖生长阶段)的迅速转换来获得高的繁殖力,最后达到较高的繁殖产量;4)繁殖分配比率均较高,分别达62.3%、51.9%、54.3%和58.3%,且其繁殖输出与植株大小呈正相关;5)萌发时间不同的植株(春、秋萌株)在大小及繁殖输出上差异显著,但繁殖分配比率差异不显著,说明由萌发时间差异引起的繁殖输出差异归根结底是由植株大小差异引起的。最后探讨了胡卢巴属这4种短命植物的生活史对策以及因萌发时间不同产生的植株在个体大小、生存力和繁殖输出等方面的差异对准噶尔荒漠干旱环境的适应意义。
姚红, 谭敦炎. 胡卢巴属4种短命植物个体大小依赖的繁殖输出与生活史对策. 植物生态学报, 2005, 29(6): 954-960. DOI: 10.17521/cjpe.2005.0124
YAO Hong, TAN Dun-Yan. SIZE-DEPENDENT REPRODUCTIVE OUTPUT AND LIFE-HISTORY STRATEGIES IN FOUR EPHEMERAL SPECIES OF TRIGONELLA. Chinese Journal of Plant Ecology, 2005, 29(6): 954-960. DOI: 10.17521/cjpe.2005.0124
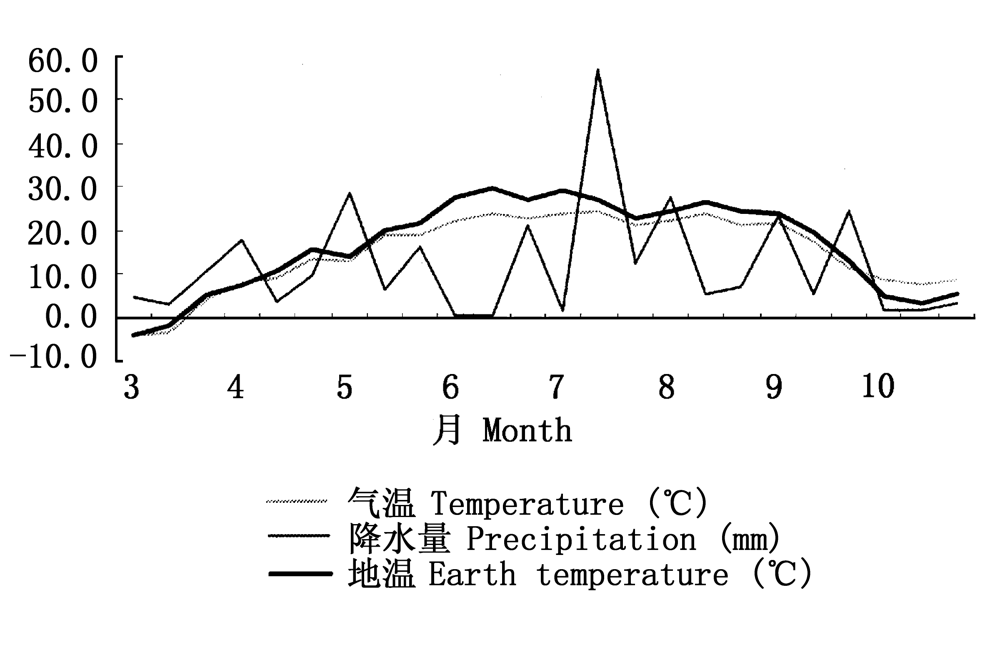
图1 观测点主要气象因子变化图(2003~2004年)
Fig.1 Main meteorological factors in observation site (2003-2004)
| 种名 Species | 植株类型 Plant type | 萌动期 Germination (日/月) (Day/Month) | 展叶期/返 青期 Seedling (日/月) (Day/Month) | 开花期 Florescence (日/月) (Day/Month) | 果熟期 Maturation (日/月) (Day/Month) | 黄枯期 Withered (日/月) (Day/Month) | 生活周期 Living circle (d) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 弯果胡卢巴 Trigonella arcuata 网脉胡卢巴 T. cancellata 直果胡卢巴 T. orthoceras 单花胡卢巴 T. monantha | 1 0 1 0 1 0 1 0 | 22/3~30/3 18/9~11/10 22/3~28/3 19/9~10/10 23/3~2/4 20/9~15/10 24/3~28/3 22/9~13/10 | 26/3~18/4 23/3~16/4 28/3~16/4 25/3~18/4 30/3~20/4 25/3~16/4 30/3~20/4 26/3~16/4 | 24/4~29/5 21/4~13/5 3/5~28/5 20/4~12/5 30/4~16/5 23/4~16/5 30/4~18/5 22/4~15/5 | 14/5~14/6 7/5~3/6 16/5~10/6 7/5~30/5 25/5~17/6 20/5~6/6 23/5~16/6 20/5~5/6 | 5/6~14/6 28/5~5/6 1/6~10/6 28/5~3/6 5/6~17/6 29/~56/6 6/6~16/6 1/6~7/6 | 53~76 232~235 55~74 231~233 63~72 233~242 60~80 234~241 | ||||
表1 春萌株与秋萌株的物候特点
Table 1 Phenological characteristics of spring and autumn germinated plants in four Trigonella species
| 种名 Species | 植株类型 Plant type | 萌动期 Germination (日/月) (Day/Month) | 展叶期/返 青期 Seedling (日/月) (Day/Month) | 开花期 Florescence (日/月) (Day/Month) | 果熟期 Maturation (日/月) (Day/Month) | 黄枯期 Withered (日/月) (Day/Month) | 生活周期 Living circle (d) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 弯果胡卢巴 Trigonella arcuata 网脉胡卢巴 T. cancellata 直果胡卢巴 T. orthoceras 单花胡卢巴 T. monantha | 1 0 1 0 1 0 1 0 | 22/3~30/3 18/9~11/10 22/3~28/3 19/9~10/10 23/3~2/4 20/9~15/10 24/3~28/3 22/9~13/10 | 26/3~18/4 23/3~16/4 28/3~16/4 25/3~18/4 30/3~20/4 25/3~16/4 30/3~20/4 26/3~16/4 | 24/4~29/5 21/4~13/5 3/5~28/5 20/4~12/5 30/4~16/5 23/4~16/5 30/4~18/5 22/4~15/5 | 14/5~14/6 7/5~3/6 16/5~10/6 7/5~30/5 25/5~17/6 20/5~6/6 23/5~16/6 20/5~5/6 | 5/6~14/6 28/5~5/6 1/6~10/6 28/5~3/6 5/6~17/6 29/~56/6 6/6~16/6 1/6~7/6 | 53~76 232~235 55~74 231~233 63~72 233~242 60~80 234~241 | ||||
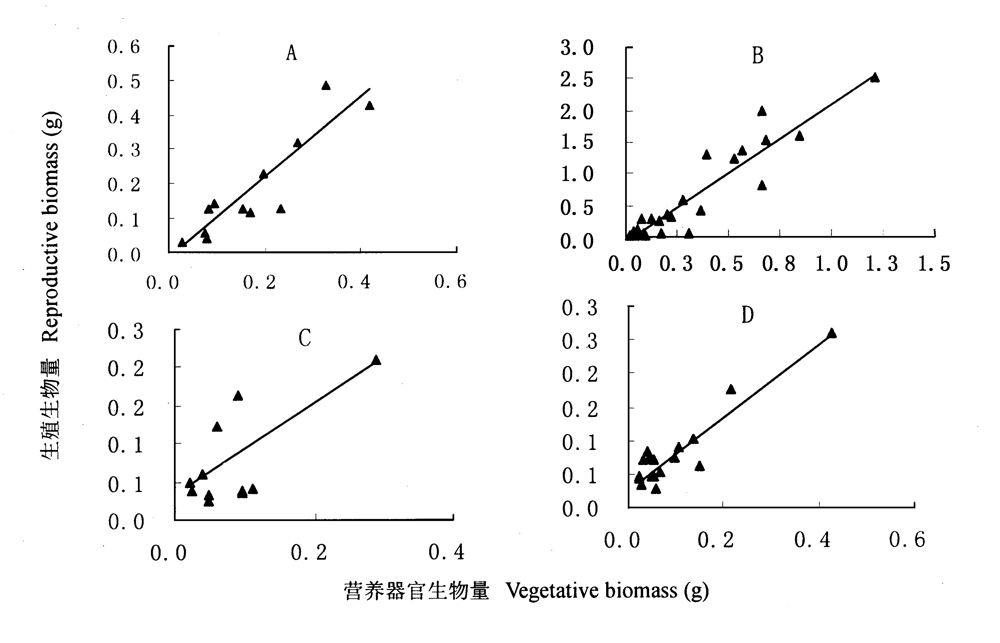
图2 繁殖输出与植株大小的相关关系 A: 弯果胡卢巴 Trigonella arcuata (r = 0.989,p<0.001) B:网脉胡卢巴 T. cancellata (r = 0.935,p<0.001) C:单花胡卢巴 T. monantha (r = 0.720, p<0.001) D: 直果胡卢巴T.orthoceras (r = 0.924,p<0.001) 在0.01水平相关性显著 Correlations are significant at the 0.01 level
Fig.2 The relationship between reproductive output and vegetative biomass
| 不同萌发时间的植株 Germination time | f | 显著性水平 p-value | |||
|---|---|---|---|---|---|
| 秋萌株 Autumn germinated plants | 春萌株 Spring germinated plants | ||||
| 弯果胡卢巴 Trigonella arcuata 网脉胡卢巴 T. cancellata 单花胡卢巴 T. monantha 直果胡卢巴 T. orthoceras | 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA | 0.436±0.115 0.641±0.222 0.745±0.114 0.569±0.066 1.126±0.164 0.615±0.037 0.127±0.019 0.142±0.011 0.548±0.031 0.103±0.014 0.135±0.025 0.536±0.024 | 0.071±0.005 0.081±0.010 0.511±0.025 0.090±0.015 0.120±0.022 0.566±0.038 0.045±0.006 0.044±0.003 0.522±0.038 0.046±0.003 0.042±0.003 0.475±0.029 | 10.143** 6.382* 4.042ns 49.502** 36.878** 0.848ns 17.072** 76.974** 0.277ns 15.358** 13.264** 2.594ns | 0.003 0.016 0.052 0.000 0.000 0.363 0.000 0.000 0.602 0.000 0.001 0.116 |
表2 4种植物秋、春萌株生物量分配的平均值±SE(n = 20)及One-Way ANOVA检验萌发时间对生物量分配的影响结果
Table 2 Biomass allocation of spring and autumn germinated plants of four species Means±SE(n=20) and the influence of germinated time on biomass allocation test by One-Way ANOVA
| 不同萌发时间的植株 Germination time | f | 显著性水平 p-value | |||
|---|---|---|---|---|---|
| 秋萌株 Autumn germinated plants | 春萌株 Spring germinated plants | ||||
| 弯果胡卢巴 Trigonella arcuata 网脉胡卢巴 T. cancellata 单花胡卢巴 T. monantha 直果胡卢巴 T. orthoceras | 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA 营养生长生物量 VB 生殖生物量 RB 生殖分配比率 RA | 0.436±0.115 0.641±0.222 0.745±0.114 0.569±0.066 1.126±0.164 0.615±0.037 0.127±0.019 0.142±0.011 0.548±0.031 0.103±0.014 0.135±0.025 0.536±0.024 | 0.071±0.005 0.081±0.010 0.511±0.025 0.090±0.015 0.120±0.022 0.566±0.038 0.045±0.006 0.044±0.003 0.522±0.038 0.046±0.003 0.042±0.003 0.475±0.029 | 10.143** 6.382* 4.042ns 49.502** 36.878** 0.848ns 17.072** 76.974** 0.277ns 15.358** 13.264** 2.594ns | 0.003 0.016 0.052 0.000 0.000 0.363 0.000 0.000 0.602 0.000 0.001 0.116 |
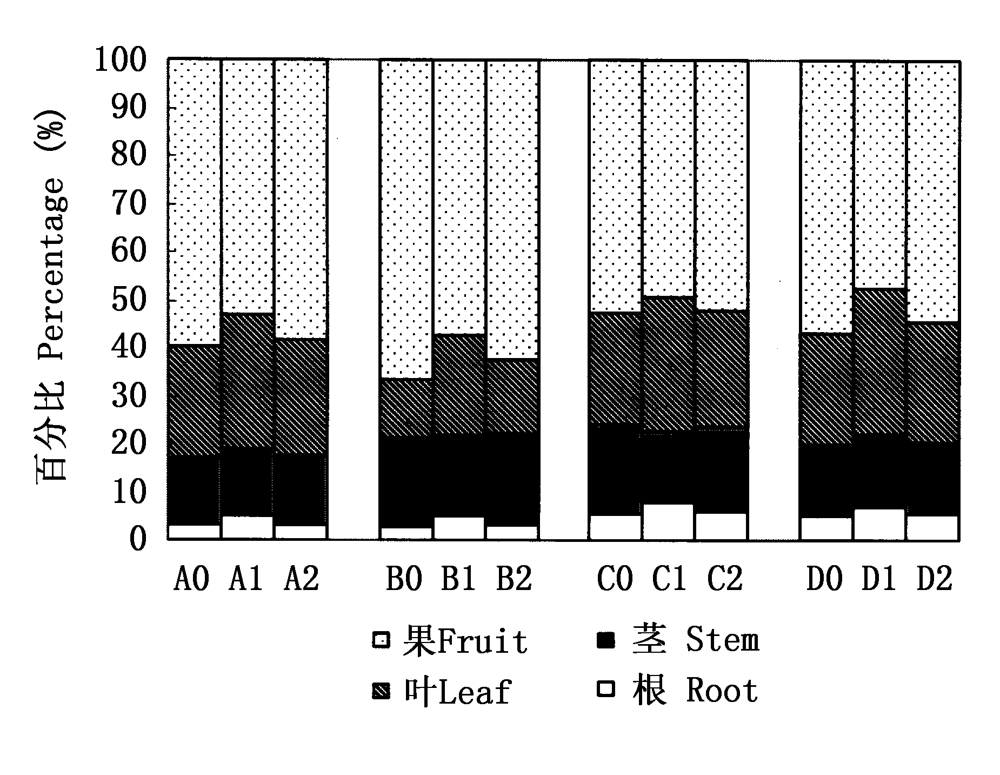
图3 4种植物的繁殖分配 A、B、C、D:同图2 See Fig.2 0:秋萌株 Autumn germinated plants 1: 春萌株 Spring germinated plants 2: 平均值 Mean
Fig.3 Reproductive allocation of four species
| [1] | Abrahamson WG, Gadgil M (1973). Growth form and reproductive effort in goldenrods (Solidago: Compositae). The American Naturalist, 107,651-661. |
| [2] | Abrahamson WG (1977). Resource allocation and growth in Impotiens capensis (Balsaminaceae) in two habitats. Bulletin of the Torrey Botanical Club, 104,160-164. |
| [3] | Abrahamson WG (1979). Pattern of resource allocation in wild flower population of fields and wood. American Journal of Botany, 66,71-79. |
| [4] | Ashmum JW, Brown RL, Pitelka LF (1985). Biomass allocation in Aster acumintus variation within and among population over 5 years. Canadian Journal of Botany, 63,2035-2043. |
| [5] | Avila-Sakar G, Krupnick GA, Stephenson AG (2001). Growth and resource allocation in Cucurbita pepo ssp. Texana: effects of fruit removal. Journal of Plant Science, 162,1089-1095. |
| [6] | Bazzaz FA, Chiariello NR, Coley PD, Pitelka LF (1987). Allocation resources to reproduction and defense. BioScience, 37,58-67. |
| [7] | Brock MA (1983). Reproductive allocation in annual and perennial species of the submerged aquatic halophyte Ruppia. Journal of Ecology, 71,811-818. |
| [8] | Fabbro T, Kørner C (2004). Altitudinal differences in flower traits and reproductive allocation. Flora, 199,70-81. |
| [9] |
Gutterman Y (1993). Seed Germination in Desert Plants. Springer-Verlag, Berlin.
DOI URL PMID |
| [10] | Harper JL (1977). The Population Biology of Plants. Academic Press, London. |
| [11] | Huang PY (黄培毅) (2002). Excused Irrigation Vegetation and its Restoration in Desert (干旱区免灌植被及其恢复). Science Press, Beijing. (in Chinese) |
| [12] |
King D, Roughgarden J (1983). Energy allocation patterns of the California grassland annuals Plantago erecta and Clarkia rubicunda. Ecology, 64,16-24.
DOI URL |
| [13] |
Klinkhamer PGL, de Jong TJ, Metz H (1997). Sex and size in cosexual plants. Trends in Ecology and Evolution, 12,260-265.
URL PMID |
| [14] | MacArthur RH, Wilson EO (1967). The Theory of Island Biogeography. Princeton University Press, Princeton. |
| [15] | Mayer AM, Poljakoff-Mayber A (1982). The Germination of Seed 3rd edn. Pergamon, Oxford. |
| [16] | Mendez M, Obeso JR (1993). Size-dependent reproductive and vegetative allocation in Arum italicum (Araceae). Canadian Journal of Botany, 71,309-314. |
| [17] | Pitelka LF (1977). Energy allocation in annual and perennial lupines ( Lupinus: Leguminosae). Ecology, 58,1055-1065. |
| [18] | Rameau C, Gouyon PH (1991). Resource allocation to growth, reproduction and survival in Gladiolus: the cost of male function. Journal of Evolutionary Biology, 14,291-307. |
| [19] | Reekie EG (1998). An explanation for size-dependent reproductive allocation in Plantago major. Canadian Journal of Botany, 76,43-50. |
| [20] | Samson DA, Werk KS (1986). Size-dependent effects in the analysis of reproductive effort in plants. The American Naturalist, 127,667-680. |
| [21] | Shem-Tov S, Gutterman D (2003). Influence of regime and photoperiod treatments on resource allocation and reproductive successes of two annuals occurring in the Negev Desert of Israel. Journal of Arid Environments, 55,123-142. |
| [22] | Susko DJ, Lovett-Doust L (2000). Plant-size and fruit-position effects on reproductive allocation in Alliaria petiolata (Brassicaceae). Canadian Journal of Botany, 78,1398-1407. |
| [23] | Thompson BK, Weiner J, Warwick SI (1991). Size-dependent reproductive output in agricultural weeds. Canadian Journal of Botany, 69,442-446. |
| [24] | Tuomi J (1983). Alternative concepts of reproductive effort, cost of reproduction and selection in life-history evolution. American Zoology, 23,25-34. |
| [25] | van Noordwijk AJ, de Jong G (1986). Acquisition and allocation of resources: their influence on variation in life history tactics. The American Naturalist, 128,137-142. |
| [26] | Vignolio OR, Fernandez ON, Maceira NO (2002). Biomass allocation to vegetative and reproductive organs in Lotus glaber and L. corniculatus (Fabaceae). Australian Journal of Botany, 50,75-82. |
| [27] | Zhang DY (张大勇) (2004). The Evolution of Life-History and Reproductive Ecology of Plant (植物生活史进化与繁殖生态学). Science Press, Beijing. (in Chinese) |
| [28] | Zhang LY (张立运) (1985). A preliminary study on the short-lived plant of Mosowan district of Xinjiang. Acta Phytoecologica et Geobotanica Sinica (植物生态学与地植物学丛刊), 9,213-221. (in Chinese with English abstract) |
| [29] | Zhao ZG (赵志刚), Du GZ (杜国祯), Ren QJ (任青吉) (2004). Size-dependent reproduction and sex allocation in five species of Ranunculaceae. Acta Phytoecologica Sinica (植物生态学报), 28,9-16. (in Chinese with English abstract) |
| [30] | Zu YG (祖元刚), Zhang WH (张文辉), Yan XF (阎秀峰), Ge S (葛颂) (1999). Conservation Biology of Endangered adenophora (濒危植物裂叶沙参保护生物学研究). Science Press, Beijing. (in Chinese) |
| [31] | Wan MW (宛敏渭), Liu XZ (刘秀珍) (1987). The Observation Methods of Phenology in China (中国物候观测方法). Science Press, Beijing. (in Chinese) |
| [32] |
Welham CVJ, Setter RA (1998). Comparison of size-dependent reproductive effort in two dandelion ( Taraxacum officinale) populations. Canadian Journal of Botany, 76,166-173.
DOI URL |
| [33] | Worley AC, Harder LD (1996). Size-dependent resource allocation and costs of reproduction in Pinguicula vulgaris. Journal of Ecology, 84,195-206. |
| [1] | 梁逸娴, 王传宽, 臧妙涵, 上官虹玉, 刘逸潇, 全先奎. 落叶松径向生长和生物量分配对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 459-468. |
| [2] | 耿雪琪, 唐亚坤, 王丽娜, 邓旭, 张泽凌, 周莹. 氮添加增加中国陆生植物生物量并降低其氮利用效率[J]. 植物生态学报, 2024, 48(2): 147-157. |
| [3] | 刘艳杰, 刘玉龙, 王传宽, 王兴昌. 东北温带森林5个羽状复叶树种叶成本-效益关系比较[J]. 植物生态学报, 2023, 47(11): 1540-1550. |
| [4] | 张玉林, 尹本丰, 陶冶, 李永刚, 周晓兵, 张元明. 早春首次降雨时间及降雨量对古尔班通古特沙漠两种短命植物形态特征与叶绿素荧光的影响[J]. 植物生态学报, 2022, 46(4): 428-439. |
| [5] | 臧永新, 马剑英, 周晓兵, 陶冶, 尹本丰, 沙亚古丽•及格尔, 张元明. 极端干旱和降水对沙垄不同坡向坡位短命植物地上生产力的影响[J]. 植物生态学报, 2022, 46(12): 1537-1550. |
| [6] | 王娇, 关欣, 张伟东, 黄苛, 朱睦楠, 杨庆朋. 杉木幼苗生物量分配格局对氮添加的响应[J]. 植物生态学报, 2021, 45(11): 1231-1240. |
| [7] | 邢磊, 段娜, 李清河, 刘成功, 李慧卿, 孙高洁. 白刺不同物候期的生物量分配规律[J]. 植物生态学报, 2020, 44(7): 763-771. |
| [8] | 曹嘉瑜, 刘建峰, 袁泉, 徐德宇, 樊海东, 陈海燕, 谭斌, 刘立斌, 叶铎, 倪健. 森林与灌丛的灌木性状揭示不同的生活策略[J]. 植物生态学报, 2020, 44(7): 715-729. |
| [9] | 张娜, 朱阳春, 李志强, 卢信, 范如芹, 刘丽珠, 童非, 陈静, 穆春生, 张振华. 淹水和干旱生境下铅对芦苇生长、生物量分配和光合作用的影响[J]. 植物生态学报, 2018, 42(2): 229-239. |
| [10] | 高景, 王金牛, 徐波, 谢雨, 贺俊东, 吴彦. 不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究[J]. 植物生态学报, 2016, 40(8): 775-787. |
| [11] | 陈青青, 李德志. 根系隔离条件下的谷子亲缘识别[J]. 植物生态学报, 2015, 39(12): 1188-1197. |
| [12] | 潘少安, 彭国全, 杨冬梅. 从叶内生物量分配策略的角度理解叶大小的优化[J]. 植物生态学报, 2015, 39(10): 971-979. |
| [13] | 肖遥,陶冶,张元明. 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征[J]. 植物生态学报, 2014, 38(9): 929-940. |
| [14] | 毛伟, 李玉霖, 崔夺, 赵学勇, 张铜会, 李玉强. 沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应[J]. 植物生态学报, 2014, 38(2): 125-133. |
| [15] | 徐波, 王金牛, 石福孙, 高景, 吴宁. 青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应[J]. 植物生态学报, 2013, 37(3): 187-196. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19