
植物生态学报 ›› 2016, Vol. 40 ›› Issue (12): 1319-1327.DOI: 10.17521/cjpe.2016.0107
郭瑞1,2,*, 周际3, 杨帆4, 李峰1, 李昊如1,2, 夏旭1,2, 刘琪1,2
出版日期:2016-12-31
发布日期:2016-12-30
通讯作者:
郭瑞
基金资助:Rui GUO1,2,*, Ji ZHOU3, Fan YANG4, Feng LI1, Hao-Ru LI1,2, Xu XIA1,2, Qi LIU1,2
Online:2016-12-31
Published:2016-12-30
Contact:
Rui GUO
摘要:
采用盆栽试验模拟干旱胁迫(土壤相对含水量40%-45%)在小麦(Triticum aestivum)拔节孕穗期胁迫12天, 测定其生长速率、光合特征及关键代谢产物含量, 以探讨干旱胁迫对拔节孕穗期小麦叶片初生及次生代谢产物的影响及其涉及的代谢途径, 讨论小麦生长代谢变化规律及应答机制。研究表明: 干旱胁迫使小麦叶片气孔受限制导致光合速率下降; 使叶绿素含量下降直接影响光系统II活性, 最终导致生长率降低。检测出的初级代谢产物组包括有机酸、氨基酸、碳水化合物、嘧啶和嘌呤等64个代谢产物, 其中29个代谢产物在干旱胁迫下发生明显的变化。主成分分析(PCA)结果显示全部样本均分布在95%的置信区间内, 两个主成分得分为64%。单因素方差分析结果表明, 干旱胁迫导致苹果酸、柠檬酸、乌头酸等参与三羧酸(TCA)循环的代谢产物消耗明显, 且引起大部分氨基酸(如脯氨酸、丝氨酸、缬氨酸)和碳水化合物(肌醇、果糖、葡萄糖)大量积累的同时转氨基代谢(天冬酰胺、谷氨酰胺和γ氨基丁酸)产物消耗, 研究证明干旱胁迫明显地促进小麦叶片的糖酵解和氨基酸合成途径, 但抑制了TCA循环和转氨基反应, 加速氨基酸代谢网络向脯氨酸合成转变过程。这些结果表明干旱胁迫引起了转氨基反应、TCA循环、糖酵解/糖异生、谷氨酸介导的脯氨酸合成, 以及嘧啶和嘌呤等代谢网络系统广泛的变化, 说明小麦在合成大量的氨基酸和碳水化合物类物质的同时也消耗了大量的能量, 暗示了糖异生到脯氨酸合成的转变。
郭瑞, 周际, 杨帆, 李峰, 李昊如, 夏旭, 刘琪. 拔节孕穗期小麦干旱胁迫下生长代谢变化规律. 植物生态学报, 2016, 40(12): 1319-1327. DOI: 10.17521/cjpe.2016.0107
Rui GUO, Ji ZHOU, Fan YANG, Feng LI, Hao-Ru LI, Xu XIA, Qi LIU. Growth metabolism of wheat under drought stress at the jointing-booting stage. Chinese Journal of Plant Ecology, 2016, 40(12): 1319-1327. DOI: 10.17521/cjpe.2016.0107
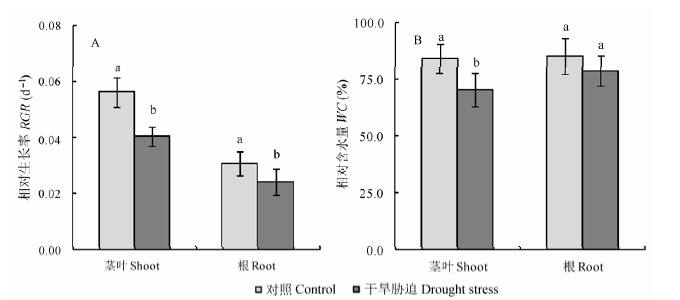
图1 干旱胁迫对冬小麦地上部分和地下部分相对生长速率(RGR) (A)和含水量(WC) (B)的影响(平均值±标准误差, n = 5)。不同小写字母表示处理间差异显著(p < 0.05, t-test)。
Fig. 1 Effects of drought stress on the relative growth rate (RGR) (A) and relative water content (WC) (B) of shoots and roots of wheat at the jointing-booting stage (mean ± SE, n = 5). Different lowercase letters indicate significant difference between treatments (p < 0.05, t-test).
| 水分处理 Water treatment | 光合参数 Photosynthetic indices | 荧光参数 Chlorophyll fluorescence | 叶绿素含量 Chlorophyll content | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 净光合速率 Pn (μmol·m-2·s-1) | 气孔导度 Gs (μmol·m-2·s-1) | PSII原初光能 转换效率 Fv/Fm | 光化学 淬灭系数qP | PSII的表观光合 电子传递速率 ETR | 叶绿素含量 Chlorophyll contents (g·kg-1 fresh mass) | 叶绿素a/ 叶绿素b Chl a/Chl b | |||
| 对照 Control | 14.98 ± 0.63a | 0.29 ± 0.01a | 0.73 ± 0.02a | 0.22 ± 0.05a | 28.28 ± 3.36a | 4.29 ± 0.88a | 2.16 ± 0.23a | ||
| 干旱胁迫 Drought stress | 3.15 ± 0.37b | 0.04 ± 0.00b | 0.31 ± 0.03b | 0.09 ± 0.00b | 12.49 ± 1.33b | 2.01 ± 0.63b | 1.31 ± 0.14b | ||
表1 干旱胁迫对冬小麦光合参数(Pn, Gs), 荧光参数(Fv/Fm, qP, ETR)和叶绿素含量(Chl a, Chl b, Car)的影响(平均值±标准误差, n = 5)
Table 1 The photosynthetic indices (Pn, Gs), chlorophyll fluorescence (Fv/Fm, qP, ETR), chlorophyll and carotenoid contents and Chl a/Chl b of wheat seedling leaves under drought stress at the jointing-booting stage (mean ± SE, n = 5)
| 水分处理 Water treatment | 光合参数 Photosynthetic indices | 荧光参数 Chlorophyll fluorescence | 叶绿素含量 Chlorophyll content | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 净光合速率 Pn (μmol·m-2·s-1) | 气孔导度 Gs (μmol·m-2·s-1) | PSII原初光能 转换效率 Fv/Fm | 光化学 淬灭系数qP | PSII的表观光合 电子传递速率 ETR | 叶绿素含量 Chlorophyll contents (g·kg-1 fresh mass) | 叶绿素a/ 叶绿素b Chl a/Chl b | |||
| 对照 Control | 14.98 ± 0.63a | 0.29 ± 0.01a | 0.73 ± 0.02a | 0.22 ± 0.05a | 28.28 ± 3.36a | 4.29 ± 0.88a | 2.16 ± 0.23a | ||
| 干旱胁迫 Drought stress | 3.15 ± 0.37b | 0.04 ± 0.00b | 0.31 ± 0.03b | 0.09 ± 0.00b | 12.49 ± 1.33b | 2.01 ± 0.63b | 1.31 ± 0.14b | ||
| 代谢通路 Metabolic Pathway | 代谢产物 Metabolite | 相对含量 Relative concentration | 变化率 Rate of change Log2 (DS/CK) | ||
|---|---|---|---|---|---|
| 对照组 Control (CK) | 干旱胁迫组 DS | ||||
| 三羧酸循环 The tricarboxylic acid cycle | 草酸 Oxalic acid | 1.85 | 1.03 | -0.84 | |
| 柠檬酸 Citric acid | 6.39 | 2.24 | -1.51* | ||
| 乌头酸 Aconitic acid | 3.37 | 1.65 | -1.03* | ||
| 异柠檬酸 Isocitric acid | 1.01 | 0.64 | -0.66 | ||
| Α-酮戊二酸 α-Ketoglutaric acid | 0.31 | 0.12 | -1.37* | ||
| 琥珀酸 Succinic acid | 1.46 | 0.99 | -0.56 | ||
| 延胡索酸 Fumaric acid | 0.59 | 0.27 | -1.13* | ||
| 苹果酸 Malic acid | 44.28 | 15.78 | -1.49* | ||
| 糖酵解 Glycolysis | 丙酮酸 Pyruvate | 0.36 | 0.42 | 0.21 | |
| 磷酸烯醇式丙酮酸 Phosphoenolpyruvate | 0.00 | 0.00 | 1.52* | ||
| 葡萄糖-6-磷酸 Fructose-6-phosphate | 0.00 | 0.01 | 1.05* | ||
| 果糖-6-磷酸 Glucose-6-phosphate | 0.05 | 0.11 | 1.09* | ||
| 葡萄糖 Glucose | 0.02 | 0.05 | 1.41* | ||
| 氨基酸类 Amino acids | 脯氨酸 Proline | 1.76 | 58.64 | 5.06* | |
| 丙氨酸 Alanine | 3.13 | 2.93 | -0.10 | ||
| 苯丙氨酸 Phenylalanine | 0.88 | 1.14 | 0.38 | ||
| 天冬氨酸 Aspartic acid | 14.38 | 6.41 | -1.17* | ||
| 天冬酰胺 Asparagine | 0.11 | 0.39 | 1.83* | ||
| 甘氨酸 Glycine | 0.26 | 0.53 | 1.03* | ||
| 丝氨酸 Serine | 5.90 | 11.90 | 1.01* | ||
| 苏氨酸 Threonine | 0.01 | 0.01 | 0.59 | ||
| 谷氨酸 Glutamate | 0.47 | 1.41 | 1.60* | ||
| 谷氨酰胺 Glutamine | 1.41 | 0.37 | -1.93* | ||
| 缬氨酸 Valine | 3.62 | 12.09 | 1.74* | ||
| 半胱氨酸 Cysteine | 0.03 | 0.05 | 0.72 | ||
| 异亮氨酸 Isoleucine | 1.61 | 7.09 | 2.14* | ||
| 亮氨酸 Leucine | 0.36 | 1.52 | 2.06* | ||
| 赖氨酸 Lysine | 0.46 | 0.48 | 0.07 | ||
| 甲硫氨酸 Methionine | 0.50 | 0.38 | -0.40 | ||
| 色氨酸 Tryptophan | 0.05 | 0.05 | 0.05 | ||
| 酪氨酸 Tyrosine | 0.01 | 0.01 | -0.12 | ||
| 鸟氨酸 Ornithine | 0.22 | 0.19 | -0.18 | ||
| 瓜氨酸 Citrulline | 0.06 | 0.10 | 0.85 | ||
| 糖类及多元醇 Sugars and polyols | 蔗糖 Sucrose | 12.77 | 6.37 | -1.00* | |
| 果糖 Fructose | 1.09 | 2.21 | 1.03* | ||
| 半乳糖 Galactose | 0.19 | 0.29 | 0.63 | ||
| 木糖 Xylose | 0.06 | 0.09 | 0.62 | ||
| 海藻糖 Trehalose | 0.02 | 0.04 | 0.97* | ||
| 肌醇 Myo-inositol | 15.06 | 30.85 | 1.03* | ||
| 甘露糖 Mannose | 21.27 | 20.53 | -0.05 | ||
| 纤维二糖 Cellobiose | 0.04 | 0.03 | -0.48 | ||
| 阿卓糖 Altrose | 1.03 | 1.34 | 0.39 | ||
| 葡庚糖 Glucoheptose | 0.38 | 0.48 | 0.35 | ||
| 夫糖 Fucose | 0.08 | 0.09 | 0.13 | ||
| 代谢通路 Metabolic Pathway | 代谢产物 Metabolite | 相对含量 Relative concentration | 变化率 Rate of change Log2 (DS/CK) | ||
| 对照组 Control (CK) | 干旱胁迫组 DS | ||||
| 糖类及多元醇 Sugars and polyols | 半乳糖苷 Galactinol | 1.57 | 1.34 | -0.23 | |
| 龙胆二糖 Gentiobiose | 0.09 | 0.20 | 1.14* | ||
| 乳糖 Lactose | 0.07 | 0.05 | -0.68 | ||
| 苏糖 Threose | 0.03 | 0.03 | 0.15 | ||
| 来苏糖 Lyxose | 20.14 | 27.01 | 0.42 | ||
| 景天庚糖 Sedoheptulose | 0.02 | 0.04 | 1.04* | ||
| 塔格糖 Tagatose | 1.74 | 2.73 | 0.65 | ||
| 核苷酸衍生物 Nucleotides derivatives | 尿嘧啶 Uridine | 0.03 | 0.03 | -0.06 | |
| 胸腺嘧啶 Thymidine | 1.82 | 0.68 | -1.42* | ||
| 鸟嘌呤 Guanosine | 0.27 | 0.17 | -0.65 | ||
| 次黄嘌呤 Hypoxanthine | 0.25 | 0.11 | -1.14* | ||
| 有机酸及其他代谢产物 Organic acids and others | γ-氨基丁酸 γ-aminobutyric acid | 28.23 | 15.43 | -0.87* | |
| 莽草酸 Shikimic acid | 11.89 | 12.61 | 0.08 | ||
| 奎尼酸 Quinic acid | 2.77 | 2.70 | -0.04 | ||
| 乙醇酸 Glyceric acid | 1.34 | 1.42 | 0.09 | ||
| 乙醇胺 Ethanolamine | 0.81 | 0.36 | -1.16* | ||
| 肉桂酸 Cinnamic acid | 0.03 | 0.04 | 0.25 | ||
| 绿原酸 Chlorogenic acid | 0.28 | 0.18 | -0.66 | ||
| 阿魏酸 Ferulic acid | 0.06 | 0.06 | 0.12 | ||
| 琥珀酸半醛 Succinate semialdehyde | 0.04 | 0.06 | 0.63 | ||
表2 拔节孕穗期干旱胁迫下小麦叶片提取物中代谢产物相对含量和变化倍数
Table 2 Relative concentration and the change of major metabolites in leaves of wheat seedlings under drought stress (DS) treatment at the jointing-booting stage
| 代谢通路 Metabolic Pathway | 代谢产物 Metabolite | 相对含量 Relative concentration | 变化率 Rate of change Log2 (DS/CK) | ||
|---|---|---|---|---|---|
| 对照组 Control (CK) | 干旱胁迫组 DS | ||||
| 三羧酸循环 The tricarboxylic acid cycle | 草酸 Oxalic acid | 1.85 | 1.03 | -0.84 | |
| 柠檬酸 Citric acid | 6.39 | 2.24 | -1.51* | ||
| 乌头酸 Aconitic acid | 3.37 | 1.65 | -1.03* | ||
| 异柠檬酸 Isocitric acid | 1.01 | 0.64 | -0.66 | ||
| Α-酮戊二酸 α-Ketoglutaric acid | 0.31 | 0.12 | -1.37* | ||
| 琥珀酸 Succinic acid | 1.46 | 0.99 | -0.56 | ||
| 延胡索酸 Fumaric acid | 0.59 | 0.27 | -1.13* | ||
| 苹果酸 Malic acid | 44.28 | 15.78 | -1.49* | ||
| 糖酵解 Glycolysis | 丙酮酸 Pyruvate | 0.36 | 0.42 | 0.21 | |
| 磷酸烯醇式丙酮酸 Phosphoenolpyruvate | 0.00 | 0.00 | 1.52* | ||
| 葡萄糖-6-磷酸 Fructose-6-phosphate | 0.00 | 0.01 | 1.05* | ||
| 果糖-6-磷酸 Glucose-6-phosphate | 0.05 | 0.11 | 1.09* | ||
| 葡萄糖 Glucose | 0.02 | 0.05 | 1.41* | ||
| 氨基酸类 Amino acids | 脯氨酸 Proline | 1.76 | 58.64 | 5.06* | |
| 丙氨酸 Alanine | 3.13 | 2.93 | -0.10 | ||
| 苯丙氨酸 Phenylalanine | 0.88 | 1.14 | 0.38 | ||
| 天冬氨酸 Aspartic acid | 14.38 | 6.41 | -1.17* | ||
| 天冬酰胺 Asparagine | 0.11 | 0.39 | 1.83* | ||
| 甘氨酸 Glycine | 0.26 | 0.53 | 1.03* | ||
| 丝氨酸 Serine | 5.90 | 11.90 | 1.01* | ||
| 苏氨酸 Threonine | 0.01 | 0.01 | 0.59 | ||
| 谷氨酸 Glutamate | 0.47 | 1.41 | 1.60* | ||
| 谷氨酰胺 Glutamine | 1.41 | 0.37 | -1.93* | ||
| 缬氨酸 Valine | 3.62 | 12.09 | 1.74* | ||
| 半胱氨酸 Cysteine | 0.03 | 0.05 | 0.72 | ||
| 异亮氨酸 Isoleucine | 1.61 | 7.09 | 2.14* | ||
| 亮氨酸 Leucine | 0.36 | 1.52 | 2.06* | ||
| 赖氨酸 Lysine | 0.46 | 0.48 | 0.07 | ||
| 甲硫氨酸 Methionine | 0.50 | 0.38 | -0.40 | ||
| 色氨酸 Tryptophan | 0.05 | 0.05 | 0.05 | ||
| 酪氨酸 Tyrosine | 0.01 | 0.01 | -0.12 | ||
| 鸟氨酸 Ornithine | 0.22 | 0.19 | -0.18 | ||
| 瓜氨酸 Citrulline | 0.06 | 0.10 | 0.85 | ||
| 糖类及多元醇 Sugars and polyols | 蔗糖 Sucrose | 12.77 | 6.37 | -1.00* | |
| 果糖 Fructose | 1.09 | 2.21 | 1.03* | ||
| 半乳糖 Galactose | 0.19 | 0.29 | 0.63 | ||
| 木糖 Xylose | 0.06 | 0.09 | 0.62 | ||
| 海藻糖 Trehalose | 0.02 | 0.04 | 0.97* | ||
| 肌醇 Myo-inositol | 15.06 | 30.85 | 1.03* | ||
| 甘露糖 Mannose | 21.27 | 20.53 | -0.05 | ||
| 纤维二糖 Cellobiose | 0.04 | 0.03 | -0.48 | ||
| 阿卓糖 Altrose | 1.03 | 1.34 | 0.39 | ||
| 葡庚糖 Glucoheptose | 0.38 | 0.48 | 0.35 | ||
| 夫糖 Fucose | 0.08 | 0.09 | 0.13 | ||
| 代谢通路 Metabolic Pathway | 代谢产物 Metabolite | 相对含量 Relative concentration | 变化率 Rate of change Log2 (DS/CK) | ||
| 对照组 Control (CK) | 干旱胁迫组 DS | ||||
| 糖类及多元醇 Sugars and polyols | 半乳糖苷 Galactinol | 1.57 | 1.34 | -0.23 | |
| 龙胆二糖 Gentiobiose | 0.09 | 0.20 | 1.14* | ||
| 乳糖 Lactose | 0.07 | 0.05 | -0.68 | ||
| 苏糖 Threose | 0.03 | 0.03 | 0.15 | ||
| 来苏糖 Lyxose | 20.14 | 27.01 | 0.42 | ||
| 景天庚糖 Sedoheptulose | 0.02 | 0.04 | 1.04* | ||
| 塔格糖 Tagatose | 1.74 | 2.73 | 0.65 | ||
| 核苷酸衍生物 Nucleotides derivatives | 尿嘧啶 Uridine | 0.03 | 0.03 | -0.06 | |
| 胸腺嘧啶 Thymidine | 1.82 | 0.68 | -1.42* | ||
| 鸟嘌呤 Guanosine | 0.27 | 0.17 | -0.65 | ||
| 次黄嘌呤 Hypoxanthine | 0.25 | 0.11 | -1.14* | ||
| 有机酸及其他代谢产物 Organic acids and others | γ-氨基丁酸 γ-aminobutyric acid | 28.23 | 15.43 | -0.87* | |
| 莽草酸 Shikimic acid | 11.89 | 12.61 | 0.08 | ||
| 奎尼酸 Quinic acid | 2.77 | 2.70 | -0.04 | ||
| 乙醇酸 Glyceric acid | 1.34 | 1.42 | 0.09 | ||
| 乙醇胺 Ethanolamine | 0.81 | 0.36 | -1.16* | ||
| 肉桂酸 Cinnamic acid | 0.03 | 0.04 | 0.25 | ||
| 绿原酸 Chlorogenic acid | 0.28 | 0.18 | -0.66 | ||
| 阿魏酸 Ferulic acid | 0.06 | 0.06 | 0.12 | ||
| 琥珀酸半醛 Succinate semialdehyde | 0.04 | 0.06 | 0.63 | ||
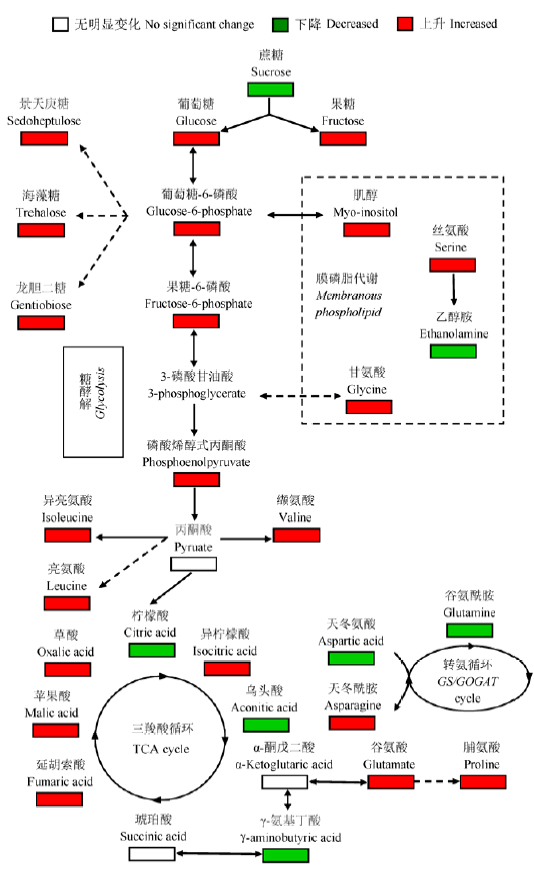
图2 干旱胁迫下小麦叶片代谢途径网络变化图。红色代表显著性增加, 绿色代表显著性降低 (p < 0.05)。
Fig. 2 Proposed changes of metabolic network of wheat leaves under drought stress. Red boxes denote significant increases while green ones denote significant decreases (p < 0.05).
| 1 | Arnon DI (1949). Copper enzymes in isolated chlorop lasts phenoloxidases inBeta vulgaris. Plant Physiology, 24, 1-15. |
| 2 | Banuelos GS, Fakra SC, Walse SS (2011). Selenium accumula- tion, distribution, and speciation in spineless prickly pear cactus: A drought- and salt-tolerant, selenium-enriched nutraceutical fruit crop for biofortified foods.Plant Physiology, 1, 315-327. |
| 3 | Bray EA (1997). Plant responses to water deficit.Trends in Plant Science, 2, 48-54. |
| 4 | Chen X (2015). Effects of Drought Stress on Growth, Yield and Quality of Different Barley Genotypes. Master degree dissertation, Zhejiang University, Hangzhou. 14.(in Chinese with English abstract)[陈雪 (2015). 干旱胁迫对不同大麦生长发育、产量和品质的影响. 硕士毕业论文. 浙江大学, 杭州. 14.] |
| 5 | Cui Q, Lewis IA, Hegeman AD, Anderson ME (2008). Metabolite identification via the madison metabolomics consortium database.Nature Biotechnology, 26, 162-164. |
| 6 | Dai H, Xiao C, Liu H, Tang H (2010). Combined NMR and LC-MS analysis reveals the metabonomic changes inSalvia miltiorrhiza Bunge induced by water depletion. Journal of Proteome Research, 9, 1460-1475. |
| 7 | Hill R, Schreiber U, Gademann R, Larkum AWD, Kühl M, Ralp PJ (2004). Spatial heterogeneity of photosynthesis and the effect of temperature-induced bleaching conditions in three species of corals.Marine Biology, 144, 633-640. |
| 8 | Hou CX, Tang ZC (1999). Function and mechanism of com- patible solutes.Plant Physiology Communications, 35, 1-7.(in Chinese with English abstract)[侯彩霞, 汤章城 (1999). 细胞相容性物质的生理功能及其作用机制. 植物生理学通讯, 35, 1-7. ] |
| 9 | Jaleel CA, Gopi R, Sankar B, Gomathinayagam M, Pann- eerselvam R (2008). Differential responses in water use efficiency in two varieties ofCatharanthus roseus under drought stress. Comptes Rendus Biologies, 331, 42-47. |
| 10 | Jiang SX, Liu DX, Pang HX, Lü JY (2014). Effects of PEG stress and recovery on activities of key enzymes involved in proline metabolism in wheat cultivars with difference in drought tolerance.Acta Botanica Boreali-Occidentalia Sinica, 34, 1581-1587.(in Chinese with English abstract)[姜淑欣, 刘党校, 庞红喜, 吕金印 (2014). PEG胁迫及复水对不同抗旱性小麦幼苗脯氨酸代谢关键酶活性的影响. 西北植物学报, 34, 1581-1587. ] |
| 11 | Kang SZ (2014). Towards water and food security in china.Chinese Journal of Eco-Agriculture, 22, 880-885.(in Chinese with English abstract)[康绍忠 (2014). 水安全与粮食安全. 中国生态农业学报, 22, 880-885. ] |
| 12 | Kingsbury RW, Epstein E (1984). Selection for salt resistant in spring wheat.Crop Science ,24, 310-315. |
| 13 | Kreps JA, Wu YJ, Chang HS, Zhu T, Wang X, Harper JF (2002). Transcriptome changes forArabidopsis in res- ponse to salt, osmotic, and cold stress. Plant Physiology, 130, 2129-2141. |
| 14 | LaRosa PC, Rhodes D, Rhodes JC, Bressan RA, Csonka LN (1991). Elevated accumulation of proline in NaCl-adapted tobacco cells is not due to altered 1-pyrroline-5- carboxylate reductase.Plant Physiology ,96, 245-250. |
| 15 | Lawlor DW, Cornic G (2002). Photosynthetic carbon assimila- tion and associated metabolism in relation to water deficits in higher plants.Plant, Cell and Environment, 25, 175-294. |
| 16 | Lisec J, Schauer N, Kopka J, Willmitzer L, Fernie AR (2006). Gas chromatography mass spectrometry-based metabolite profiling in plants.Nature Protocol, 1, 387-396. |
| 17 | Lü LH, Hu YK, Li YM (2006). Dynamics of proline accumulation in winter wheat under different water stress.Acta Agriculturae Boreali-Sinica, 21, 75-78.(in Chinese with English abstract)[吕丽华, 胡玉昆, 李雁鸣 (2006). 水分胁迫下不同抗旱性冬小麦脯氨酸积累动态. 华北农学报, 2l, 75-78.] |
| 18 | Wang JY, Zhu SG, Xu CF (2002). Biochemistry. 3rd edn. Shengcai Education, Beijing. [王镜岩, 朱圣庚, 许长法 (2002). 生物化学. 第三版. 圣才教育出版社, 北京.] |
| 19 | Wang L, Liu Y, Li DQ (2012). Drought stress signal transduction and regulation mechanism in plants.Biotechnology Bulletin, 10, 1-7.(in Chinese with English abstract)[王丽, 刘洋, 李德全 (2012). 植物干旱胁迫信号转导及其调控机制研究进展. 生物技术通报, 10, 1-7.] |
| 20 | Wu Y, He L, Hu WH (2009). Advances in regulatory metabolic networks in drought stress responses.Hubei Agricultural Sciences, 48, 1504-1509.(in Chinese with English abstract)[吴杨,贺俐,胡文海 (2009). 植物干旱胁迫下的调控代谢网络研究进展. 湖北农业科学, 48, 1504-1509. ] |
| 21 | Wu YC, Zhou SL, Wang ZM (2004). Review on genetic im- provement in root related to drought-resistance in wheat.Journal of Triticeae Crops, 24, 101-104.(in Chinese with English abstract)[吴永成, 周顺利, 王志敏 (2004). 小麦 |
| 22 | 与抗旱性有关的根系遗传改良研究进展. 麦类作物学报, 24, 101-104. ] |
| 23 | Xu MJ, Liu GR, Yang XJ, Wang LJ (2002). Study on drought induced protein of winter wheat varieties. Journal Publishing Department of Agricultural University of Hebei, 2, 11-15.(in Chinese) [徐民俊, 刘桂茹, 杨学举, 王丽军 (2002). 冬小麦品种干旱诱导蛋白的研究. 河北农业大学学报, 2, 11-15.] |
| 24 | Yang C, Shi D, Wang D (2008). Comparative effects of salt stress and alkali stress on growth, osmotic adjustment and ionic balance of an alkali resistant halophyteSuaeda glauca(Bge.). Plant Growth Regular, 56, 179-190. |
| 25 | Yang WP, Shan CJ, Hu XQ, Li J (2008). Effects of soil drought on carbon metabolism of winter wheat during jointing stage.Journal of Henan Agricultural Sciences, 9, 20-26.(in Chinese with English abstract)[杨文平, 单长卷, 胡喜巧, 李杰 (2008). 土壤干旱对冬小麦拔节期叶片碳代谢的影响. 河南农业科学, 9, 20-26.] |
| 26 | Zhang RH, Zheng YJ, Ma GS, Zhang XH, Lu HD, Shi JT, Xue JQ (2011). Effects of drought stress on photosynthetic traits and protective enzyme activity in maize seeding.Acta Ecologica Sinica, 31, 1303-1311.(in Chinese with English abstract)[张仁和,郑友军,马国胜,张兴华,路海东,史俊通,薛吉全 (2011). 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响. 生态学报, 31, 1303-1311.] |
| 27 | Zhao LL, Xu QJ, Jiang Y, Li YH (2008). The mitogen- activated protein kinase signal transduction in plant cell under biotic and abiotic stress conditions.Plant Phy- siology Communications, 1, 196-174.(in Chinese with English abstract)[赵琳琳, 徐启江, 姜勇, 李玉花 (2008). 生物和非生物胁迫下的植物细胞中丝裂原活化蛋白激酶(MAPK)信号转导. 植物生理学通讯, 1, 196-174. ] |
| 28 | Zhao YJ, Weng BQ, Wang YX, Xu GZ (2009). Physiological and ecological responses of plants to drought stress and its research progress.Fujian Science and Technology of Rice and Wheat, 27, 46-50.(in Chinese with English abstract)[赵雅静, 翁伯琦, 王义祥, 徐国忠 (2009). 植物对干旱胁迫的生理生态响应及其研究进展. 福建稻麦科技, 27, 46-50.] |
| [1] | 陈图强, 徐贵青, 刘深思, 李彦. 干旱胁迫下梭梭水力性状调整与非结构性碳水化合物动态[J]. 植物生态学报, 2023, 47(10): 1407-1421. |
| [2] | 周洁, 杨晓东, 王雅芸, 隆彦昕, 王妍, 李浡睿, 孙启兴, 孙楠. 梭梭和骆驼刺对干旱的适应策略差异[J]. 植物生态学报, 2022, 46(9): 1064-1076. |
| [3] | 熊淑萍, 曹文博, 曹锐, 张志勇, 付新露, 徐赛俊, 潘虎强, 王小纯, 马新明. 水平结构配置对冬小麦冠层垂直结构、微环境及产量的影响[J]. 植物生态学报, 2022, 46(2): 188-196. |
| [4] | 刘丽燕, 冯锦霞, 刘文鑫, 万贤崇. 干旱胁迫对转PtPIP2;8基因84K杨苗木光合、生长和根系结构的影响[J]. 植物生态学报, 2020, 44(6): 677-686. |
| [5] | 徐丽娇, 郝志鹏, 谢伟, 李芳, 陈保冬. 丛枝菌根真菌根外菌丝跨膜H +和Ca 2+流对干旱胁迫的响应[J]. 植物生态学报, 2018, 42(7): 764-773. |
| [6] | 王曦,胡红玲,胡庭兴,张城浩,王鑫,刘丹. 干旱胁迫对桢楠幼树渗透调节与活性氧代谢的影响及施氮的缓解效应[J]. 植物生态学报, 2018, 42(2): 240-251. |
| [7] | 罗丹丹, 王传宽, 金鹰. 植物水分调节对策: 等水与非等水行为[J]. 植物生态学报, 2017, 41(9): 1020-1032. |
| [8] | 郭瑞, 周际, 杨帆, 李峰. 小麦根系在碱胁迫下的生理代谢反应[J]. 植物生态学报, 2017, 41(6): 683-692. |
| [9] | 徐静馨, 郑有飞, 麦博儒, 赵辉, 储仲芳, 黄积庆, 袁月. 基于涡度相关法的麦田O3干沉降及不同沉降通道分配的特征[J]. 植物生态学报, 2017, 41(6): 670-682. |
| [10] | 高林, 王晓菲, 顾行发, 田庆久, 焦俊男, 王培燕, 李丹. 植冠下土壤类型差异对遥感估算冬小麦叶面积指数的影响[J]. 植物生态学报, 2017, 41(12): 1273-1288. |
| [11] | 岑宇, 刘美珍. 凝结水对干旱胁迫下羊草和冰草生理生态特征及叶片形态的影响[J]. 植物生态学报, 2017, 41(11): 1199-1207. |
| [12] | 郑成岩, 邓艾兴, LATIFMANESHHojatollah, 宋振伟, 张俊, 王利, 张卫建. 增温对青藏高原冬小麦干物质积累转运及氮吸收利用的影响[J]. 植物生态学报, 2017, 41(10): 1060-1068. |
| [13] | 郭瑞, 李峰, 周际, 李昊儒, 夏旭, 刘琪. 亚麻响应盐、碱胁迫的生理特征[J]. 植物生态学报, 2016, 40(1): 69-79. |
| [14] | 安东升, 曹娟, 黄小华, 周娟, 窦美安. 基于Lake模型的叶绿素荧光参数在甘蔗苗期抗旱性研究中的应用[J]. 植物生态学报, 2015, 39(4): 398-406. |
| [15] | 金皖豫, 李铭, 何杨辉, 杜正刚, 邵钧炯, 张国栋, 周灵燕, 周旭辉. 不同施氮水平对冬小麦生长期土壤呼吸的影响[J]. 植物生态学报, 2015, 39(3): 249-257. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19