
植物生态学报 ›› 2018, Vol. 42 ›› Issue (5): 526-538.DOI: 10.17521/cjpe.2017.0305
所属专题: 全球变化与生态系统; 生态遥感及应用; 青藏高原植物生态学:生态系统生态学
周彤1,曹入尹2,王少鹏1,陈晋3,唐艳鸿1,*( )
)
收稿日期:2017-11-21
修回日期:2018-02-11
出版日期:2018-05-20
发布日期:2018-07-20
通讯作者:
唐艳鸿
基金资助:
ZHOU Tong1,CAO Ru-Yin2,WANG Shao-Peng1,CHEN Jin3,TANG Yan-Hong1,*()
Received:2017-11-21
Revised:2018-02-11
Online:2018-05-20
Published:2018-07-20
Contact:
Yan-Hong TANG
Supported by:摘要:
长期以来, 气候与植物物候关系的研究大多基于线性模型, 但植被物候对气候变化的响应可能是非线性的。该文利用非线性模型——生存分析模型来分析时间序列中过去事件(气候因子)对目的变量(物候)的作用: 用生存分析模型分析了春季气温和降水量对内蒙古草地、青藏高原草甸和欧洲地区木本植物返青期的影响。其中, 内蒙古与青藏高原的物候信息来自遥感数据, 欧洲地区物候信息为实测数据。蒙特卡洛方法用于拟合模型参数。结果表明: 生存分析模型适合对上述不同研究对象的物候影响因素进行分析, 并能模拟非线性效应; 在内蒙古草地, 春季气温和降水对春季返青有很大的影响, 而青藏高原草甸和欧洲地区木本植物返青期对春季降水响应较小, 对春季气温变化的响应随Holdridge干燥度指数上升而下降; 在预测返青期时发现: 春季平均气温提高1 ℃会导致上述地区返青期提前1-6天; 而春季气温与降水的增加会导致返青期发生明显的非线性变化, 这种非线性效应无法基于线性模型模拟出来。结果说明生存分析模型既能用于分析不同尺度下植物物候与气候的关系, 也能用于模型预测, 尤其适合探讨大幅度气候变化对物候的非线性影响。
周彤, 曹入尹, 王少鹏, 陈晋, 唐艳鸿. 中国草地和欧洲木本植物返青期对气温和降水变化的响应: 基于生存分析的研究. 植物生态学报, 2018, 42(5): 526-538. DOI: 10.17521/cjpe.2017.0305
ZHOU Tong, CAO Ru-Yin, WANG Shao-Peng, CHEN Jin, TANG Yan-Hong. Responses of green-up dates of grasslands in China and woody plants in Europe to air temperature and precipitation: Empirical evidences based on survival analysis. Chinese Journal of Plant Ecology, 2018, 42(5): 526-538. DOI: 10.17521/cjpe.2017.0305
| 属 Genus | 物种 Species | 植株数 Plant No. |
|---|---|---|
| 七叶树属 Aesculus | 欧洲七叶树 Aesculus hippocastanum | 31 |
| 桤木属 Alnus | Alnus glutinosa | 4 |
| 桦木属 Betula | 垂枝桦 Betula pendula | 19 |
| 山毛榉属 Fagus | Fagus sylvatica | 16 |
| 栎属 Quercus | 夏栎 Quercus robur | 16 |
| 梣属 Fraxinus | 欧梣 Fraxinus excelsior | 9 |
| 茶藨子属 Ribes | Ribes grossularia | 12 |
表1 欧洲研究区木本植物物种
Table 1 List of woody plant species in European database
| 属 Genus | 物种 Species | 植株数 Plant No. |
|---|---|---|
| 七叶树属 Aesculus | 欧洲七叶树 Aesculus hippocastanum | 31 |
| 桤木属 Alnus | Alnus glutinosa | 4 |
| 桦木属 Betula | 垂枝桦 Betula pendula | 19 |
| 山毛榉属 Fagus | Fagus sylvatica | 16 |
| 栎属 Quercus | 夏栎 Quercus robur | 16 |
| 梣属 Fraxinus | 欧梣 Fraxinus excelsior | 9 |
| 茶藨子属 Ribes | Ribes grossularia | 12 |
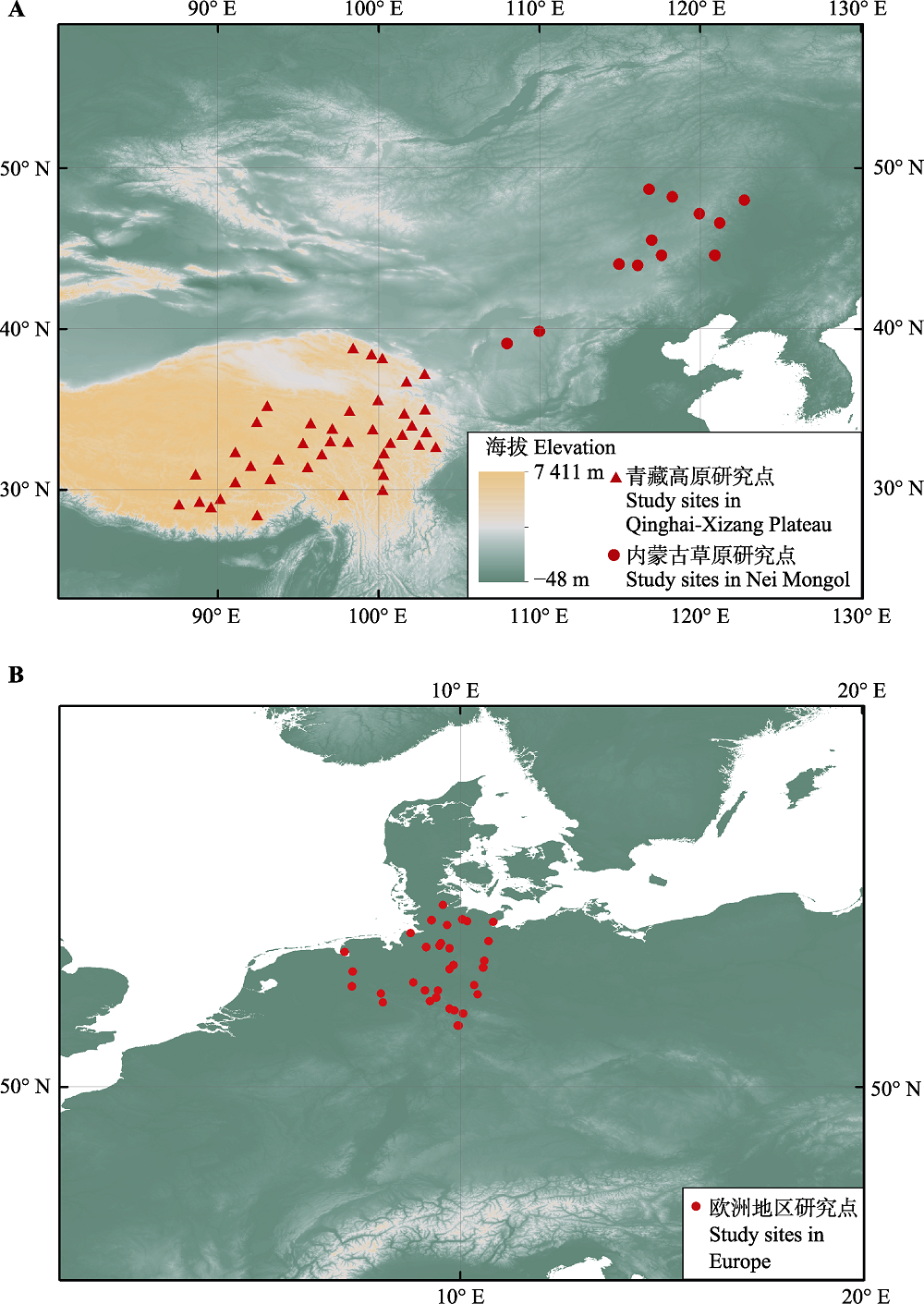
图1 中国与欧洲的研究点分布。 A图中三角形表示位于青藏高原的气象站点, 圆点表示位于内蒙古的气象站点。B图中圆点表示欧洲地区木本植物物候观测点。
Fig. 1 Spatial distribution of study sites in China (A) and Europe (B). The triangles/dots in (A) are the distributions of meteorological stations in Qinghai-Xizang Plateau and in Nei Mongol, respectively. The points in (B) indicate the locations of phenological observation sites for woody plants in Europe.
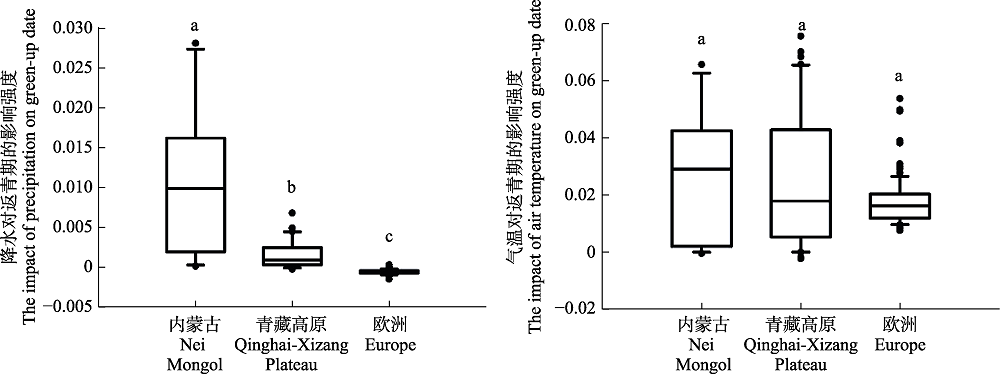
图2 春季降水与气温对内蒙古草原、青藏高原草甸和欧洲地区木本植物返青期的影响。每一方框的上下横线分别表示样本统计量的四分位值即75%和25%的位点, 框内中央横线是中位数, 上下端短横线分别是统计量的90%和10%的位点, 圆点表示统计分布的特异最大或最小值。
Fig. 2 The changes in model coefficients of green-up date with spring precipitation and air temperature for the grasslands in Nei Mongol, meadows in Qinghai-Xizang Plateau, and woody plants in Europe. In these boxplots, the top and bottom values of the bars indicate the 25th and 75th percentiles, respectively; the black line within the box indicates the median; whiskers below and above the box indicate the 10th and 90th percentiles; and points indicate outliers. Letters on top of the whiskers are the results of an analysis of variance: different letters indicate statistically significant difference between the mean values (p < 0.01, t-test).
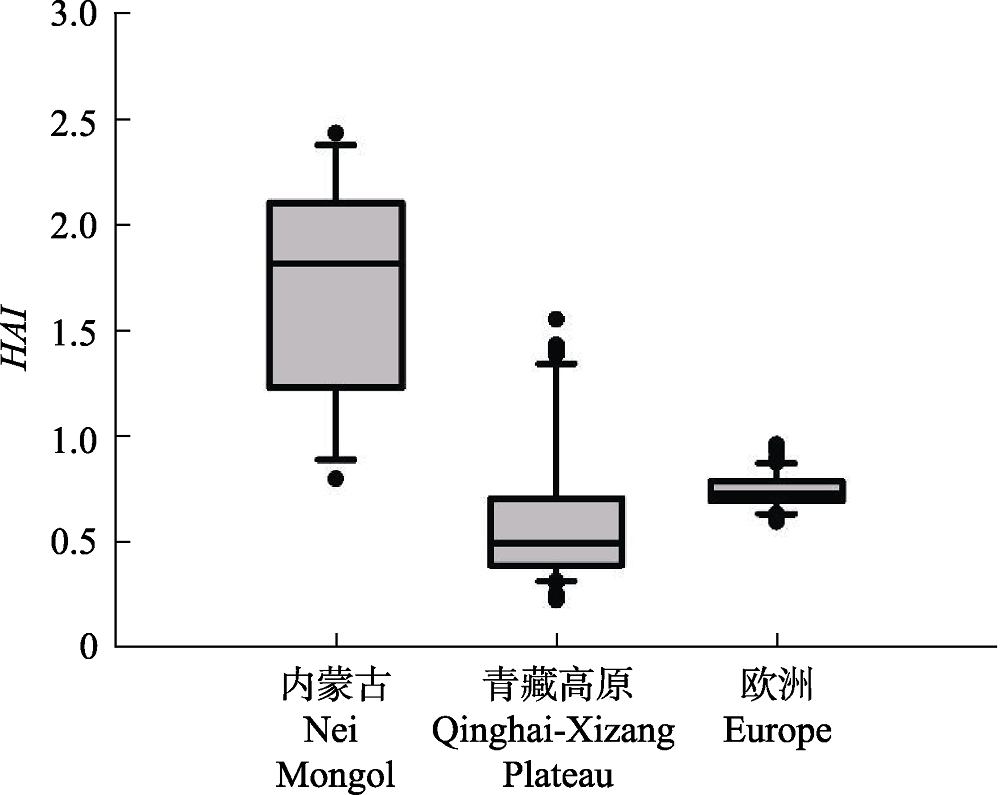
图3 内蒙古草原、青藏高原草甸和欧洲地区木本植物的Holdridge干燥度指数(HAI)的分布统计特征。统计变量的说明参见图2。
Fig. 3 Boxplots of the Holdridge aridity index (HAI) for the grasslands of Nei Mongol, meadows in Qinghai-Xizang Plateau, and woody plants in Europe. See Fig. 2 for explanations of the symbols.
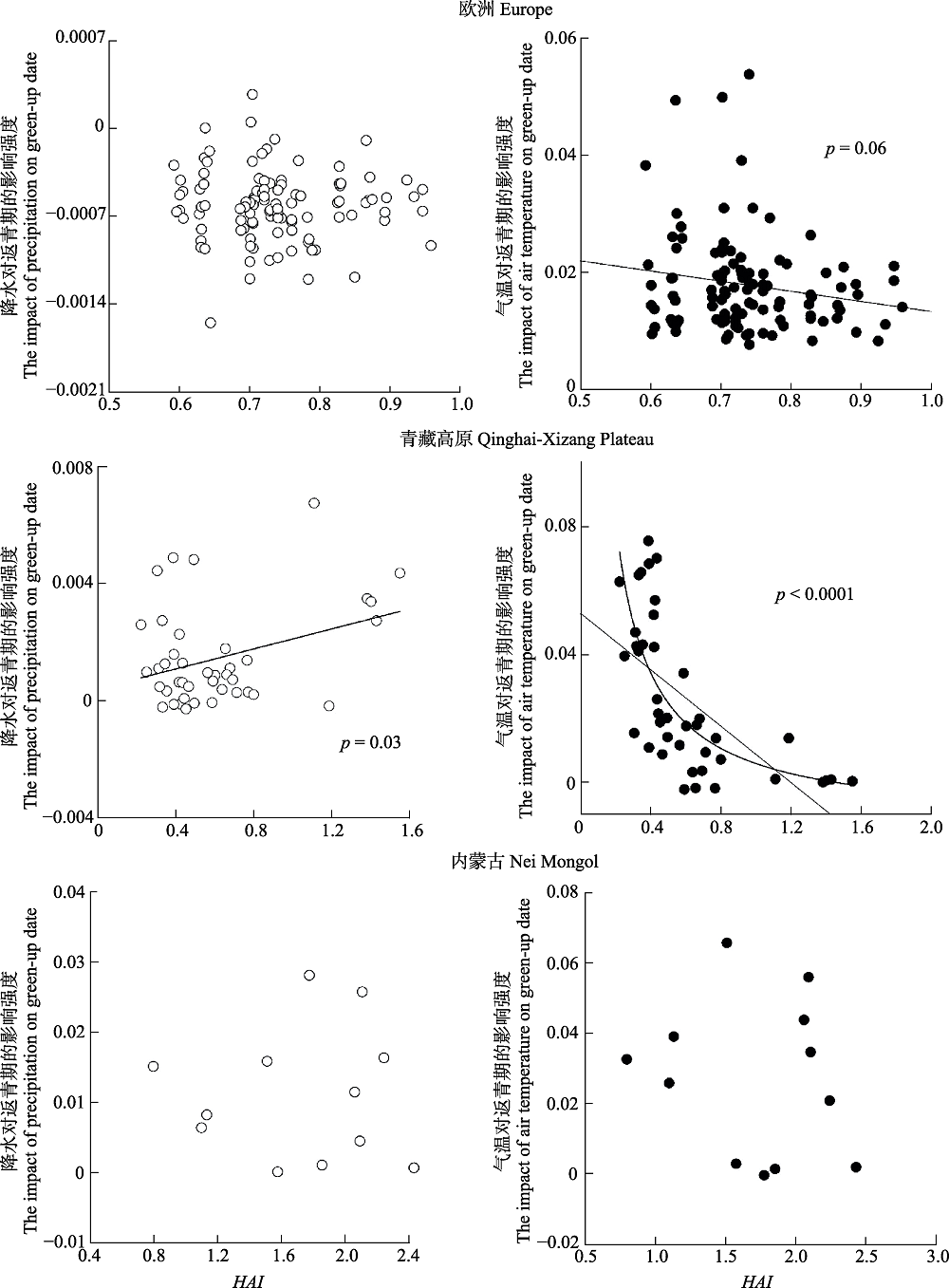
图4 内蒙古草原、青藏高原草甸和欧洲地区木本植物降水与气温对返青期的影响强度与Holdridge干燥度指数(HAI)的关系。
Fig. 4 The changes in the model coefficient of precipitation and temperature with the Holdridge aridity index (HAI) for the grasslands of Nei Mongol,meadows in Qinghai-Xizang Plateau, and woody plants in Europe.
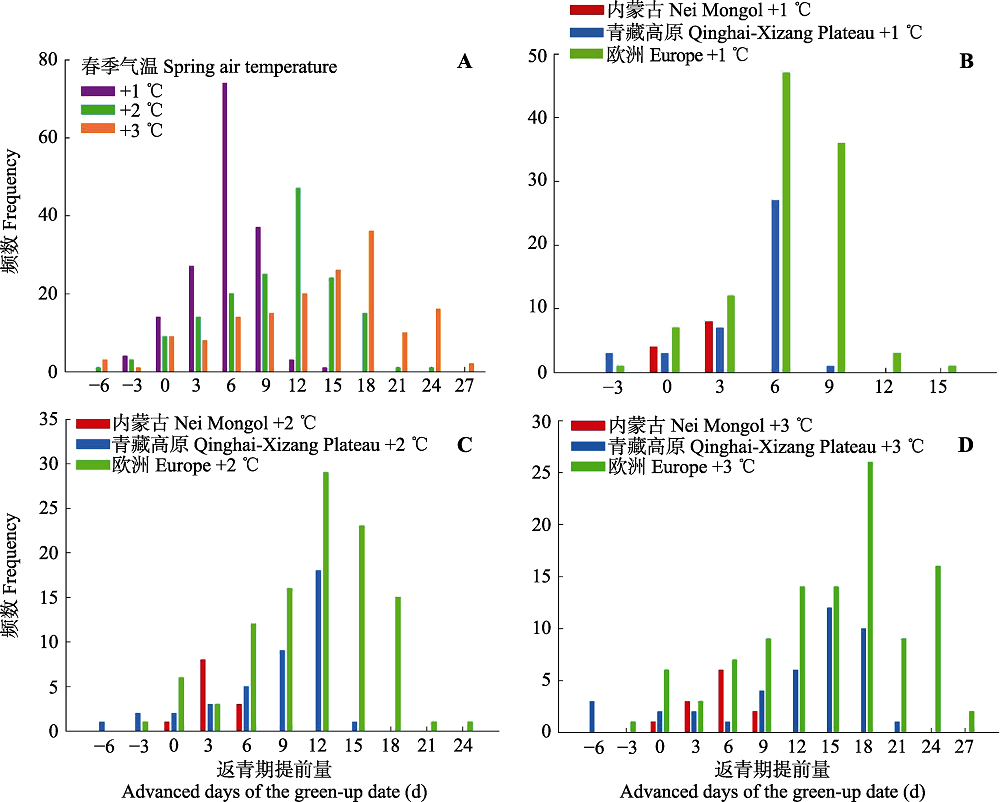
图5 在春季气温上升的3种情境下(+1、+2、+3 ℃)所研究生态系统返青期的变化。负值为返青期延后, 正值为返青期提前。A, 升温1、2和3 ℃情况下所有研究点返青期变化量的频数。B、C、D分别为升温1、2 和3 ℃时各个生态系统返青期变化量的频数。
Fig. 5 Predicted changes of green-up date under the three scenarios of elevated spring-temperature at 1, 2 and 3 °C for the target areas. Negative and positive dates indicate the delayed and advanced days of the green-up date, respectively. A, Predicted changes under all the three temperature scenarios. B, C, D, Predicted changes for each target area with the temperature increase of 1, 2 and 3 °C, respectively.
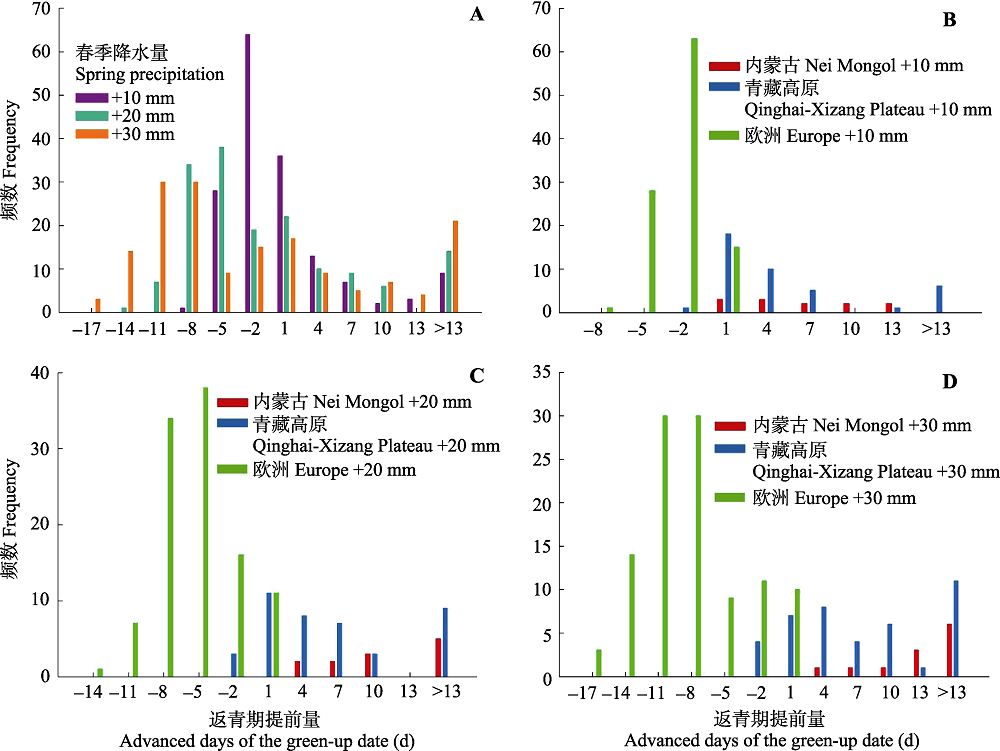
图6 在3种春季降水增加的情景下(+10、+20、+30 mm)所研究生态系统返青期的变化情况。负值表示返青期延后, 正值表示返青期提前。A, 增水10、20和30 mm情况下所有研究点返青期变化量的频数。B、C、D分别为增水10、20和30 mm时各个生态系统返青期变化量的频数。
Fig. 6 Predicted changes of green-up date under the three scenarios of elevated spring-precipitation at 10, 20 and 30 mm among each target area. Negative and positive dates indicate the delayed and advanced days of the green-up date, respectively. A, Predicted changes for all the three precipitation scenarios. B, C, D, Predicted changes for each target area with the precipitation increase at 10, 20 and 30 mm, respectively.
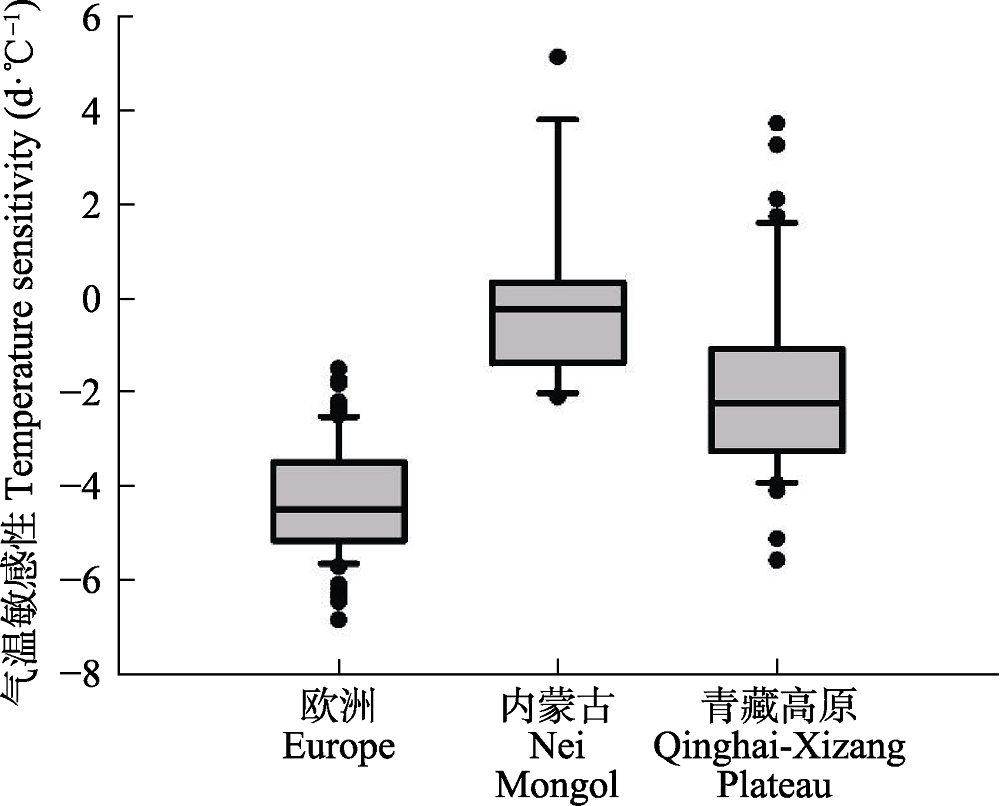
图7 利用线性回归模型计算的3个地区返青期气温敏感性。统计变量的说明参见图2。
Fig. 7 Boxplots of temperature sensitivity of green-up date from linear regression models for the three cases of the study. See Fig. 2 for explanations of the symbols.
| 升温情景 Warming scenarios (℃) | 内蒙古 Nei Mongol | 青藏高原 Qinghai-Xizang Plateau | 欧洲 Europe | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 生存分析法 Survival analysis | 线性回归法 Linear analysis | p | 生存分析法 Survival analysis | 线性回归法 Linear analysis | p | 生存分析法 Survival analysis | 线性回归法 Linear analysis | p | |
| + 1 | 0.92 | 0.13 | 0.04 | 3.56 | 1.87 | < 0.001 | 5.66 | 4.47 | < 0.001 |
| + 2 | 2.33 | 0.26 | 0.01 | 7.29 | 3.74 | < 0.001 | 10.5 | 8.94 | < 0.001 |
| + 3 | 4.17 | 0.39 | < 0.01 | 10.88 | 5.61 | < 0.001 | 14.06 | 13.41 | < 0.001 |
表2 升温1 、2 和3 ℃下内蒙古、青藏高原和欧洲地区返青期提前天数的平均值(从3月1日起每天比2009年升温1、2和3 ℃)
Table 2 The mean advanced days of green-up date for the two grasslands in Nei Mongol and the Qinghai-Xizang Plateau, and woody plants in Europe under three warming scenarios: elevated spring temperature at +1, +2 and +3 °C for each day as compared with the days from March 1st, 2009.
| 升温情景 Warming scenarios (℃) | 内蒙古 Nei Mongol | 青藏高原 Qinghai-Xizang Plateau | 欧洲 Europe | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 生存分析法 Survival analysis | 线性回归法 Linear analysis | p | 生存分析法 Survival analysis | 线性回归法 Linear analysis | p | 生存分析法 Survival analysis | 线性回归法 Linear analysis | p | |
| + 1 | 0.92 | 0.13 | 0.04 | 3.56 | 1.87 | < 0.001 | 5.66 | 4.47 | < 0.001 |
| + 2 | 2.33 | 0.26 | 0.01 | 7.29 | 3.74 | < 0.001 | 10.5 | 8.94 | < 0.001 |
| + 3 | 4.17 | 0.39 | < 0.01 | 10.88 | 5.61 | < 0.001 | 14.06 | 13.41 | < 0.001 |
| [1] |
Allen JM, Terres MA, Katsuki T, Iwamoto K, Kobori H, Higuchi H, Primack RB, Wilson AM, Gelfand A, Silander JA ( 2014). Modeling daily flowering probabilities: Expected impact of climate change on Japanese cherry phenology. Global Change Biology, 20, 1251-1263.
DOI URL PMID |
| [2] |
Bradley AV, Gerard FF, Barbier N, Weedon GP, Anderson LO, Huntingford C, Aragão LEOC, Zelazowski P, Arai E ( 2011). Relationships between phenology, radiation and precipitation in the Amazon region. Global Change Biology, 17, 2245-2260.
DOI URL |
| [3] |
Chen J, Jönsson P, Tamura M, Gu Z, Matsushita B, Eklundh L ( 2004). A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter. Remote Sensing of Environment, 91, 332-344.
DOI URL |
| [4] |
Chen J, Rao Y, Shen M, Wang C, Zhou Y, Ma L, Tang Y, Yang X ( 2016). A simple method for detecting phenological change from time series of vegetation idexindex. IEEE Transactions on Geoscience & Remote Sensing, 54, 3436-3449.
DOI URL |
| [5] | Chen W, Yu SZ ( 1997). Comparison of the Cox Model and log-linear models in analysing survival data. Chinese Journal of Health Statistics, 14, 18-20. |
| [ 陈文, 俞顺章 ( 1997). Cox回归模型与对数线性回归模型在生存分析中应用的比较. 中国卫生统计, 14, 18-20.] | |
| [6] |
Cox DR ( 1972). Regression models and life-tables. Biometrical Journal, 34, 187.
DOI URL |
| [7] |
Diez JM, Ibanez I, Silander Jr JA, Primack R, Higuchi H, Kobori H, Sen A, James TY ( 2014). Beyond seasonal climate: Statistical estimation of phenological responses to weather. Ecological Applications, 24, 1793-1802.
DOI URL PMID |
| [8] |
Ding M, Zhang Y, Sun X, Liu L, Wang Z, Bai W ( 2013). Spatiotemporal variation in alpine grassland phenology in the Qinghai-Tibetan Plateau from 1999 to 2009. Chinese Science Bulletin, 58, 396-405.
DOI URL |
| [9] |
Forkel M, Migliavacca M, Thonicke K, Reichstein M, Schaphoff S, Weber U, Carvalhais N ( 2015). Codominant water control on global interannual variability and trends in land surface phenology and greenness. Global Change Biology, 21, 3414-3435.
DOI URL PMID |
| [10] |
Fu YH, Zhao H, Piao S, Peaucelle M, Peng S, Zhou G, Ciais P, Huang M, Menzel A, Peñuelas J, Song Y, Vitasse Y, Zeng Z, Janssens IA ( 2015). Declining global warming effects on the phenology of spring leaf unfolding. Nature, 526, 104-107.
DOI URL PMID |
| [11] |
Grace J, Allen SJ, Wilson C ( 1989). Climate and the meristem temperatures of plant communities near the tree-line. Oecologia, 79, 198-204.
DOI URL |
| [12] |
Güsewell S, Furrer R, Gehrig R, Pietragalla B ( 2017). Changes in temperature sensitivity of spring phenology with recent climate warming in Switzerland are related to shifts of the preseason. Global Change Biology, 23, 5189-5202.
DOI URL PMID |
| [13] |
Haylock MR, Hofstra N, Klein Tank AMG, Klok EJ, Jones PD, New M ( 2008). A European daily high-resolution gridded data set of surface temperature and precipitation for 1950-2006. Journal of Geophysical Research, 113, D20119. DOI: 10.1029/2008JD010201.
DOI URL |
| [14] |
Iler AM, Hoye TT, Inouye DW, Schmidt NM ( 2013). Long-term trends mask variation in the direction and magnitude of short-term phenological shifts. American Journal of Botany, 100, 1398-1406.
DOI URL PMID |
| [15] |
Jochner S, Sparks TH, Laube J, Menzel A ( 2016). Can we detect a nonlinear response to temperature in European plant phenology? International Journal of Biometeorology, 60, 1551-1561.
DOI URL PMID |
| [16] | Li XZ, Han GD, Guo CY ( 2013). Impacts of climate change on dominant pasture growing season in central Inner Mongolia. Acta Ecologica Sinica, 33, 4146-4155. |
| [ 李夏子, 韩国栋, 郭春燕 ( 2013). 气候变化对内蒙古中部草原优势牧草生长季的影响. 生态学报, 33, 4146-4155.] | |
| [17] | Lian JS, Jiang HS ( 1995). The ecological meaning and its identification of the Logistic population growth model. Supplement to the Journal of Sun Yatsen University, ( 3), 160-164. |
| [ 练健生, 江海声 ( 1995). Logistic种群增长模型参数的生态学意义及其辨识──南湾野生猕猴种群的应用. 中山大学学报论丛, ( 3), 160-164.] | |
| [18] |
Mazer SJ, Travers SE, Cook BI, Davies TJ, Bolmgren K, Kraft NJB, Salamin N, Inouye DW ( 2013). Flowering date of taxonomic families predicts phenological sensitivity to temperature: Implications for forecasting the effects of climate change on unstudied taxa. American Journal of Botany, 100, 1381-1397.
DOI URL PMID |
| [19] | Meng M, Ni J, Zhang ZG ( 2004). Aridity index and its applications in geo-ecological study. Acta Phytoecologica Sinica, 28, 853-861. |
| [ 孟猛, 倪健, 张治国 ( 2004). 地理生态学的干燥度指数及其应用评述. 植物生态学报, 28, 853-861.] | |
| [20] |
Menzel A, Sparks TH, Estrella N, Koch E, Aasa A, Ahas R, Alm-Kübler K, Bissolli P, Braslavská O, Briede A, Chmielewski FM, Crepinsek Z, Curnel Y, Dahl Å, Defila C, Donnelly A, Filella Y, Jatczak K, Måge F, Mestre A, Nordli Ø, Peñuelas J, Pirinen P, Remišová V, Scheifinger H, Striz M, Susnik A, van Vliet AJH, Wielgolaski F, Zach S, Zust A ( 2006). European phenological response to climate change matches the warming pattern. Global Change Biology, 12, 1969-1976.
DOI URL |
| [21] |
Niu JM ( 2001). Impacts prediction of climatic change on distribution and production of grassland in Inner Mongolia. Acta Agrestia Sinica, 9, 277-282.
DOI URL |
|
[ 牛建明 ( 2001). 气候变化对内蒙古草原分布和生产力影响的预测研究. 草地学报, 9, 277-282.]
DOI URL |
|
| [22] | Richard P, Liu HY, Yin Y ( 2011). Conservation of biodiversity under impact of human activities: Vegetation evolution in Central Europe and its implication, Chinese Journal of Ecology, 30, 584-588. |
| [ Richard P, 刘鸿雁, 印轶 ( 2011). 人类活动影响下的生物多样性保护: 中欧的植被演化及其启示. 生态学杂志, 30, 584-588.] | |
| [23] |
Sadras VO, Moran MA ( 2013). Nonlinear effects of elevated temperature on grapevine phenology. Agricultural and Forest Meteorology, 173, 107-115.
DOI URL |
| [24] | Sarmiento G, Monasterio M ( 1983). Life Forms and Phenology: Tropical Savannas. Elsevier,Amsterdam. 79-104. |
| [25] |
Shen M, Cong N, Cao R ( 2015 a). Temperature sensitivity as an explanation of the latitudinal pattern of green-up date trend in Northern Hemisphere vegetation during 1982-2008. International Journal of Climatology, 35, 3707-3712.
DOI URL |
| [26] |
Shen M, Piao S, Cong N, Zhang G, Jassens IA ( 2015 b). Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Global Change Biology, 21, 3647-3656.
DOI URL PMID |
| [27] |
Shen M, Tang Y, Chen J, Yang X, Wang C, Cui X, Yang Y, Han L, Li L, Du J, Zhang G, Cong N ( 2014). Earlier-?season vegetation has greater temperature sensitivity of spring phenology in Northern Hemisphere. POLS ONE, 9, e88178. DOI: 10.1371/journal.pone.0088178.
DOI URL PMID |
| [28] |
Shen M, Tang Y, Chen J, Zhu X, Zheng Y ( 2011). Influences of temperature and precipitation before the growing season on spring phenology in grasslands of the central and eastern Qinghai-Tibetan Plateau. Agricultural and Forest Meteorology, 151, 1711-1722.
DOI URL |
| [29] |
Sun HL, Zheng D, Yao TD, Zhang YL ( 2012). Protection and construction of the national ecological security shelter zone on Tibetan Plateau. Acta Geographica Sinca, 67, 3-12.
DOI URL |
|
[ 孙鸿烈, 郑度, 姚檀栋, 张镱锂 ( 2012). 青藏高原国家生态安全屏障保护与建设. 地理学报, 67, 3-12.]
DOI URL |
|
| [30] | The Editorial Committee of Vegetation Map of China, Chinese Academy of Sciences ( 2001). 1:1000000 Vegetation Atlas of China. Secience Press, Beijing . |
| [ 中国科学院中国植被图编辑委员会 ( 2001). 1:1000000中国植被图集. 科学出版社, 北京] | |
| [31] |
Vitasse Y, Hoch G, Randin CF, Lenz A, Kollas C, Scheepens JF, Koerner C ( 2013). Elevational adaptation and plasticity in seedling phenology of temperate deciduous tree species. Oecologia, 171, 663-678.
DOI URL |
| [32] |
Wang C, Chen J, Wu J, Tang Y, Shi P, Black TA, Zhu K ( 2017). A snow-free vegetation index for improved monitoring of vegetation spring green-up date in deciduous ecosystems. Remote Sensing of Environment, 196, 1-12.
DOI URL |
| [33] |
Wang SP, Meng FD, Duan JC, Wang YF, Cui XY, Piao SL, Niu HS, Xu GP, Luo CY, Zhang ZH, Zhu XX, Shen MG, Li YN, Du MY, Tang YH, Zhao XQ, Ciais P, Kimball B, Penuelas J, Janssens IA, Cui SJ, Zhao L, Zhang FW ( 2014). Asymmetric sensitivity of first flowering date to warming and cooling in alpine plants. Ecology, 95, 3387-3398.
DOI URL |
| [34] |
White MA, de Beurs KM, Didan K, Inouye DW, Richardson AD, Jensen OP, O’Keefe J, Zhang G, Nemani RR, van Leeuwen WJD, Brown JF, de Wit A, Schaepman M, Lin X, Dettinger M, Bailey AS, Kimball J, Schwartz MD, Baldocchi DD, Lee JT, Lauenroth WK ( 2009). Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982-2006. Global Change Biology, 15, 2335-2359.
DOI URL |
| [35] |
Xie Y, Ahmed KF, Allen JM, Wilson AM, Silander JA ( 2015). Green-up of deciduous forest communities of northeastern North America in response to climate variation and climate change. Landscape Ecology, 30, 109-123.
DOI URL |
| [36] |
Yu H, Luedeling E, Xu J ( 2010). Winter and spring warming result in delayed spring phenology on the Tibetan Plateau. Proceedings of the National Academy of Sciences of the United States of America, 107, 22151-22156.
DOI URL PMID |
| [37] | Zhou GS, Zhang XS ( 1996). Study on climate-vegetation classification for global change in China. Acta Botanica Sinica, 38, 8-17. |
| [ 周广胜, 张新时 ( 1996). 全球变化的中国气候-植被分类研究. 植物学报, 38, 8-17.] |
| [1] | 陈科宇 邢森 唐玉 孙佳慧 任世杰 张静 纪宝明. 不同草地型土壤丛枝菌根真菌群落特征及其驱动因素[J]. 植物生态学报, 2024, 48(5): 660-674. |
| [2] | 陈以恒 玉素甫江·如素力 阿卜杜热合曼·吾斯曼. 2001-2020年天山新疆段草地植被覆盖度时空变化及驱动因素分析[J]. 植物生态学报, 2024, 48(5): 561-576. |
| [3] | 白皓然 侯盟 刘艳杰. 少花蒺藜草入侵与干旱对羊草草原生产力的影响机制[J]. 植物生态学报, 2024, 48(5): 577-589. |
| [4] | 张计深, 史新杰, 刘宇诺, 吴阳, 彭守璋. 气候变化下中国潜在自然植被生态系统碳储量动态[J]. 植物生态学报, 2024, 48(4): 428-444. |
| [5] | 臧妙涵, 王传宽, 梁逸娴, 刘逸潇, 上官虹玉, 全先奎. 基于纬度移栽的落叶松叶、枝、根生态化学计量特征对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 469-482. |
| [6] | 梁逸娴, 王传宽, 臧妙涵, 上官虹玉, 刘逸潇, 全先奎. 落叶松径向生长和生物量分配对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 459-468. |
| [7] | 吴茹茹, 刘美珍, 谷仙, 常馨月, 郭立月, 蒋高明, 祁如意. 气候变化对巨柏适宜生境分布的潜在影响和预测[J]. 植物生态学报, 2024, 48(4): 445-458. |
| [8] | 杨宇萌, 来全, 刘心怡. 气候变化和人类活动对内蒙古植被总初级生产力的定量影响[J]. 植物生态学报, 2024, 48(3): 306-316. |
| [9] | 张启, 程雪寒, 王树芝. 北京西山老龄树记载的森林干扰历史[J]. 植物生态学报, 2024, 48(3): 341-348. |
| [10] | 茹雅倩, 薛建国, 葛萍, 李钰霖, 李东旭, 韩鹏, 杨天润, 储伟, 陈章, 张晓琳, 李昂, 黄建辉. 高频轮牧对典型草原生产生态效果的影响[J]. 植物生态学报, 2024, 48(2): 171-179. |
| [11] | 索南吉, 李博文, 吕汪汪, 王文颖, 拉本, 陆徐伟, 宋扎磋, 陈程浩, 苗琪, 孙芳慧, 汪诗平. 增温增水情景下钉柱委陵菜物候序列的变化及其抗冻性[J]. 植物生态学报, 2024, 48(2): 158-170. |
| [12] | 陈颖洁, 房凯, 秦书琪, 郭彦军, 杨元合. 内蒙古温带草地土壤有机碳组分含量和分解速率的空间格局及其影响因素[J]. 植物生态学报, 2023, 47(9): 1245-1255. |
| [13] | 赵艳超, 陈立同. 土壤养分对青藏高原高寒草地生物量响应增温的调节作用[J]. 植物生态学报, 2023, 47(8): 1071-1081. |
| [14] | 李兆光, 文高, 和桂青, 徐天才, 和琼姬, 侯志江, 李燕, 薛润光. 滇西北藜麦氮磷钾生态化学计量特征的物候期动态[J]. 植物生态学报, 2023, 47(5): 724-732. |
| [15] | 任培鑫, 李鹏, 彭长辉, 周晓路, 杨铭霞. 洞庭湖流域植被光合物候的时空变化及其对气候变化的响应[J]. 植物生态学报, 2023, 47(3): 319-330. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19