
植物生态学报 ›› 2018, Vol. 42 ›› Issue (5): 595-608.DOI: 10.17521/cjpe.2018.0021
• 研究论文 • 上一篇
辜翔1,张仕吉1,刘兆丹1,李雷达1,陈金磊1,王留芳1,方晰1,2,3,*( )
)
收稿日期:2018-01-15
修回日期:2018-05-09
出版日期:2018-05-20
发布日期:2018-07-20
通讯作者:
方晰
基金资助:
GU Xiang1,ZHANG Shi-Ji1,LIU Zhao-Dan1,LI Lei-Da1,CHEN Jin-Lei1,WANG Liu-Fang1,FANG Xi1,2,3,*()
Received:2018-01-15
Revised:2018-05-09
Online:2018-05-20
Published:2018-07-20
Contact:
Xi FANG
Supported by:摘要:
为揭示植被恢复对土壤有机碳(SOC)库的影响机制, 采用空间代替时间的方法, 以湘中丘陵区地域相邻、环境条件基本一致的檵木(Loropetalum chinense)-南烛(Vaccinium bracteatum)-杜鹃(Rhododendron simsii)灌草丛(LVR)、檵木-杉木(Cunninghamia lanceolata)-白栎(Quercus fabri)灌木林(LCQ)、马尾松(Pinus massoniana)-柯(又名石栎)(Lithocarpus glaber)-檵木针阔混交林(PLL)和柯-红淡比(Cleyera japonica)-青冈(Cyclobalanopsis glauca)常绿阔叶林(LAG)作为一个恢复演替序列, 设置固定样地, 采集0-10、10-20、20-30、30-40 cm土层土壤样品, 测定不同恢复阶段SOC含量(CSOC)和SOC密度(DSOC), 通过主成分分析方法和逐步回归分析方法分析影响CSOC、DSOC变化的主要因子。结果表明: (1)各土层CSOC、DSOC随着植被恢复呈增加趋势, 且LAG显著高于其他3个恢复阶段。LAG 0-40 cm土层CSOC分别比LVR、LCQ、PLL增加12.5、9.3和4.7 g·kg -1, 分别提高了248.5%、113.1%和58.5%; DSOC分别增加67.1、46.1和32.5 t C·hm -2, 分别提高了182.0%、79.7%和45.6%。(2) CSOC、DSOC与群落植物多样性指数、群落总生物量、地上部分生物量、根系生物量、凋落物层现存量、凋落物层全氮(N)含量、凋落物层全磷(P)含量、土壤全磷(TP)、土壤有效磷(AP)含量、土壤C/N(除CSOC外)、C/P、N/P、<0.002 mm黏粒百分含量呈显著或极显著正相关关系, 与凋落物层C/N (除DSOC外)、凋落物层C/P、土壤pH值和土壤容重呈极显著负相关关系, 表明CSOC、DSOC随着植被恢复的变化受到植被因子和土壤因子诸多因子的影响。其中, 土壤C/P、土壤pH值和凋落物层C/P对CSOC、DSOC影响显著; 此外, <0.002 mm黏粒百分含量也显著影响着DSOC, 而土壤C/P对CSOC和DSOC影响最显著。植被恢复过程中, 凋落物层C/P和土壤C/P、pH值、质地的变化是影响SOC库变化的重要因素。
辜翔, 张仕吉, 刘兆丹, 李雷达, 陈金磊, 王留芳, 方晰. 中亚热带植被恢复对土壤有机碳含量、碳密度的影响. 植物生态学报, 2018, 42(5): 595-608. DOI: 10.17521/cjpe.2018.0021
GU Xiang, ZHANG Shi-Ji, LIU Zhao-Dan, LI Lei-Da, CHEN Jin-Lei, WANG Liu-Fang, FANG Xi. Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of China. Chinese Journal of Plant Ecology, 2018, 42(5): 595-608. DOI: 10.17521/cjpe.2018.0021
| 恢复阶段 Restoration stage | 优势植物 Dominant plants | 组成比例 Composition proportion (%) | 木本植物密度 Density of woody plants (trees·hm-2) | 多样性 指数 Diversity index | 重要值Important value (%) | 平均胸径 Average DBH (cm) | 平均树高 Average tree height (m) | 海拔 Elevation (m) | 坡向 Slope aspect | 坡度 Slope |
|---|---|---|---|---|---|---|---|---|---|---|
| 檵木-南烛- 杜鹃灌草丛 LVR | 檵木 Loropetalum chinense | 34.48 | 18125 | 0.87 | 27.46 | - | 0.85 (0.3-1.8) | 120-131 | 东南 Southeast | 18° |
| 南烛 Vaccinium bracteatum | 21.55 | 18.96 | ||||||||
| 杜鹃 Rhododendron simsii | 12.07 | 14.00 | ||||||||
| 白栎 Quercus fabri | 7.76 | 10.66 | ||||||||
| 板栗 Castanea mollissima | 5.17 | 2.72 | ||||||||
| 其他(8种) Others (8 species) | 18.97 | 26.20 | ||||||||
| 檵木-杉木- 白栎灌木林 LCQ | 檵木 Loropetalum chinense | 17.47 | 7633 | 1.06 | 22.02 | 2.74 (1.0-9.8) | 3.37 (1.5-6.5) | 120-135 | 西北 Northwest | 22° |
| 杉木 Cunninghamia lanceolata | 14.85 | 13.16 | ||||||||
| 白栎 Quercus fabri | 12.66 | 11.10 | ||||||||
| 南烛 Vaccinium bracteatum. | 12.66 | 9.26 | ||||||||
| 木姜子 Litsea spp. | 11.35 | 10.13 | ||||||||
| 其他(16种) Others (16 species) | 31.01 | 34.43 | ||||||||
| 马尾松-柯- 檵木针阔 混交林 PLL | 马尾松 Pinus massoniana | 39.69 | 17629 | 1.91 | 45.34 | 5.70 (1.0-28.0) | 6.54 (1.5-20.0) | 135-160 | 西南 Southwest | 20° |
| 柯 Lithocarpus glaber | 25.52 | 13.87 | ||||||||
| 檵木 Loropetalum chinense | 11.06 | 7.43 | ||||||||
| 红淡比 Cleyera japonica | 3.59 | 4.11 | ||||||||
| 连蕊茶 Camellia cuspidata | 3.11 | 4.02 | ||||||||
| 其他(22种) Others (22 species) | 17.03 | 25.23 | ||||||||
| 柯-红淡比- 青冈常绿 阔叶林 LAG | 柯 Lithocarpus glaber | 38.78 | 19970 | 2.29 | 25.75 | 5.63 (1.0-40.0) | 5.75 (1.5-20.0) | 200-260 | 东南 Southeast | 22° |
| 红淡比 Cleyera japonica | 18.70 | 11.05 | ||||||||
| 青冈 Cyclobalanopsis glauca | 5.82 | 8.90 | ||||||||
| 杉木 Cunninghamia lanceolata | 5.36 | 6.14 | ||||||||
| 格药柃 Eurya muricata Dunn | 5.06 | 5.73 | ||||||||
| 其他(31种) Others (31 species) | 26.28 | 42.44 |
表1 不同植被恢复阶段样地的基本特征
Table 1 Stand characteristics at different stages during vegetation restoration
| 恢复阶段 Restoration stage | 优势植物 Dominant plants | 组成比例 Composition proportion (%) | 木本植物密度 Density of woody plants (trees·hm-2) | 多样性 指数 Diversity index | 重要值Important value (%) | 平均胸径 Average DBH (cm) | 平均树高 Average tree height (m) | 海拔 Elevation (m) | 坡向 Slope aspect | 坡度 Slope |
|---|---|---|---|---|---|---|---|---|---|---|
| 檵木-南烛- 杜鹃灌草丛 LVR | 檵木 Loropetalum chinense | 34.48 | 18125 | 0.87 | 27.46 | - | 0.85 (0.3-1.8) | 120-131 | 东南 Southeast | 18° |
| 南烛 Vaccinium bracteatum | 21.55 | 18.96 | ||||||||
| 杜鹃 Rhododendron simsii | 12.07 | 14.00 | ||||||||
| 白栎 Quercus fabri | 7.76 | 10.66 | ||||||||
| 板栗 Castanea mollissima | 5.17 | 2.72 | ||||||||
| 其他(8种) Others (8 species) | 18.97 | 26.20 | ||||||||
| 檵木-杉木- 白栎灌木林 LCQ | 檵木 Loropetalum chinense | 17.47 | 7633 | 1.06 | 22.02 | 2.74 (1.0-9.8) | 3.37 (1.5-6.5) | 120-135 | 西北 Northwest | 22° |
| 杉木 Cunninghamia lanceolata | 14.85 | 13.16 | ||||||||
| 白栎 Quercus fabri | 12.66 | 11.10 | ||||||||
| 南烛 Vaccinium bracteatum. | 12.66 | 9.26 | ||||||||
| 木姜子 Litsea spp. | 11.35 | 10.13 | ||||||||
| 其他(16种) Others (16 species) | 31.01 | 34.43 | ||||||||
| 马尾松-柯- 檵木针阔 混交林 PLL | 马尾松 Pinus massoniana | 39.69 | 17629 | 1.91 | 45.34 | 5.70 (1.0-28.0) | 6.54 (1.5-20.0) | 135-160 | 西南 Southwest | 20° |
| 柯 Lithocarpus glaber | 25.52 | 13.87 | ||||||||
| 檵木 Loropetalum chinense | 11.06 | 7.43 | ||||||||
| 红淡比 Cleyera japonica | 3.59 | 4.11 | ||||||||
| 连蕊茶 Camellia cuspidata | 3.11 | 4.02 | ||||||||
| 其他(22种) Others (22 species) | 17.03 | 25.23 | ||||||||
| 柯-红淡比- 青冈常绿 阔叶林 LAG | 柯 Lithocarpus glaber | 38.78 | 19970 | 2.29 | 25.75 | 5.63 (1.0-40.0) | 5.75 (1.5-20.0) | 200-260 | 东南 Southeast | 22° |
| 红淡比 Cleyera japonica | 18.70 | 11.05 | ||||||||
| 青冈 Cyclobalanopsis glauca | 5.82 | 8.90 | ||||||||
| 杉木 Cunninghamia lanceolata | 5.36 | 6.14 | ||||||||
| 格药柃 Eurya muricata Dunn | 5.06 | 5.73 | ||||||||
| 其他(31种) Others (31 species) | 26.28 | 42.44 |
| 恢复阶段 Restoration stage | 群落总生物量 Community total biomass (kg·hm-2) | 地上部分 生物量 Aboveground biomass (kg·hm-2) | 根系生物量 Root biomass (kg·hm-2) | 凋落物层 现存量 Existing biomass in litter layer (kg·hm-2) | 凋落物层 C含量 C concentration in litter layer (g·kg-1) | 凋落物层 N含量 N concentration in litter layer (g·kg-1) | 凋落物层 P含量 P concentration in litter layer (g·kg-1) | 凋落物层 C/N比 C/N ratio in litter layer | 凋落物层 C/P比 C/P ratio in litter layer | 凋落物层N/P比 N/P ratio in litter layer |
|---|---|---|---|---|---|---|---|---|---|---|
| LVQ | 5 185.4 ± 2 702.5a | 2 244.5 ± 1 153.9a | 2 029.6 ± 1 515.7a | 911.4 ± 653. 9a | 315.8 ± 25.3ac | 9.84 ± 0.3a | 0.30 ± 0.02a | 35.8 ± 2.6a | 1532.2 ± 95.2a | 41.5 ± 2.9a |
| LCQ | 2 0591.5 ± 9 728.9b | 10 346.7 ± 7 143.2b | 5 443.0 ± 3 467.5a | 4 801.8 ± 1 030.2b | 277.2 ± 33.3a | 11.2 ± 0.9a | 0.34 ± 0.03ab | 26.0 ± 0.2b | 914.2 ± 69.4b | 35.7 ± 3.6a |
| PLL | 129 112.7 ± 19 713.5c | 107 692.6 ± 16 361.6c | 16 332.9 ± 3 047.6b | 5 087.2 ± 1 246.1b | 424.1 ± 8.6b | 11.5 ± 0.5a | 0.33 ± 0.05ab | 38.0 ± 1.6a | 1389.7 ± 227.9ab | 36.6 ± 4.4a |
| LAG | 148 975.4 ± 43 906.4c | 120 695.2 ± 13 647.7c | 24 400.4 ± 5 124.8c | 3 879.8 ± 1 171.5b | 332.3 ± 53.9c | 14.0 ± 0.2b | 0.38 ± 0.04b | 23.9 ± 3.5b | 953.1 ± 44.6b | 40.3 ± 4.0a |
表2 不同植被恢复阶段的群落生物量和凋落物层养分特征(平均值±标准偏差)
Table 2 Plant community biomass and nutrient characteristics of litter layer at different stages during vegetation restoration (mean ± SD)
| 恢复阶段 Restoration stage | 群落总生物量 Community total biomass (kg·hm-2) | 地上部分 生物量 Aboveground biomass (kg·hm-2) | 根系生物量 Root biomass (kg·hm-2) | 凋落物层 现存量 Existing biomass in litter layer (kg·hm-2) | 凋落物层 C含量 C concentration in litter layer (g·kg-1) | 凋落物层 N含量 N concentration in litter layer (g·kg-1) | 凋落物层 P含量 P concentration in litter layer (g·kg-1) | 凋落物层 C/N比 C/N ratio in litter layer | 凋落物层 C/P比 C/P ratio in litter layer | 凋落物层N/P比 N/P ratio in litter layer |
|---|---|---|---|---|---|---|---|---|---|---|
| LVQ | 5 185.4 ± 2 702.5a | 2 244.5 ± 1 153.9a | 2 029.6 ± 1 515.7a | 911.4 ± 653. 9a | 315.8 ± 25.3ac | 9.84 ± 0.3a | 0.30 ± 0.02a | 35.8 ± 2.6a | 1532.2 ± 95.2a | 41.5 ± 2.9a |
| LCQ | 2 0591.5 ± 9 728.9b | 10 346.7 ± 7 143.2b | 5 443.0 ± 3 467.5a | 4 801.8 ± 1 030.2b | 277.2 ± 33.3a | 11.2 ± 0.9a | 0.34 ± 0.03ab | 26.0 ± 0.2b | 914.2 ± 69.4b | 35.7 ± 3.6a |
| PLL | 129 112.7 ± 19 713.5c | 107 692.6 ± 16 361.6c | 16 332.9 ± 3 047.6b | 5 087.2 ± 1 246.1b | 424.1 ± 8.6b | 11.5 ± 0.5a | 0.33 ± 0.05ab | 38.0 ± 1.6a | 1389.7 ± 227.9ab | 36.6 ± 4.4a |
| LAG | 148 975.4 ± 43 906.4c | 120 695.2 ± 13 647.7c | 24 400.4 ± 5 124.8c | 3 879.8 ± 1 171.5b | 332.3 ± 53.9c | 14.0 ± 0.2b | 0.38 ± 0.04b | 23.9 ± 3.5b | 953.1 ± 44.6b | 40.3 ± 4.0a |
| 恢复 阶段 Restoration stage | 土层 深度 Soil layer (cm) | 容重 Bulk density (g·cm-3) | 0.05-2 mm 砂粒百分含量 Soil sand (0.05-2 mm) percentage (%) | 0.002-0.05 mm粉粒百分含量 Soil silt (0.002- 0.05 mm) percentage (%) | <0.002 mm 黏粒百分含量 Soil clay (<0.002 mm) percentage (%) | pH | TP (g·kg-1) | AP (mg·kg-1) | C/N | C/P | N/P |
|---|---|---|---|---|---|---|---|---|---|---|---|
| LVR | 0-10 | 1.4 ± 0.2Aa | 43.6 ± 4.8Aa | 44.7 ± 6.4Aa | 11.7 ± 4.6Aa | 4.5 ± 0.2Aa | 0.17 ± 0.16Aa | 2.1 ± 0.7Aa | 22.4 ± 12.5Aa | 109.1 ± 44.7Aa | 5.3 ± 2.4Aa |
| 10-20 | 1.5 ± 0.1Aab | 39.5 ± 4.1Aa | 57.0 ± 5.2Ab | 3.5 ± 1.1Ab | 4.7 ± 0.2Ab | 0.12 ± 0.03Aab | 1.4 ± 0.6Ab | 20.0 ± 9.3Aab | 59.5 ± 51.4Ab | 3.3 ± 3.0Ab | |
| 20-30 | 1.5 ± 0.1Aab | 42.0 ± 5.6Aa | 56.5 ± 6.2Ab | 2.7 ± 0.9Ab | 4.8 ± 0.2Ab | 0.11 ± 0.04Ab | 1.3 ± 0.5Ab | 17.7 ± 9.5Aab | 42.9 ± 40.9Ab | 2.5 ± 2.2Ab | |
| 30-40 | 1.5 ± 0.1Ab | 43.1 ± 5.6Aa | 54.4 ± 6.1ACb | 2.5 ± 0.6Ab | 5.0 ± 0.2Ac | 0.11 ± 0.04Ab | 1.2 ± 0.5Ab | 13.7 ± 7.7ABb | 30.5 ± 37.4Ab | 2.5 ± 2.5Ab | |
| LCQ | 0-10 | 1.4 ± 0.1Ab | 67.9 ± 2.1Ba | 21.6 ± 2.5Ba | 10.6 ± 2.4Aa | 4.8 ± 0.2Ba | 0.12 ± 0.03Aa | 2.6 ± 1.1ABa | 17.7 ± 6.8Aa | 164.4 ± 45.5Ba | 10.0 ± 3.1Ba |
| 10-20 | 1.6 ± 0.1Ab | 60.1 ± 11.1Bb | 27.2 ± 9.1Bb | 12.9 ± 5.0Ba | 4.9 ± 0.2ABab | 0.10 ± 0.03Ab | 1.9 ± 0.6Bab | 16.3 ± 4.6ABab | 90.2 ± 29.9Bb | 6.0 ± 2.8Bb | |
| 20-30 | 1.6 ± 0.1Ab | 63.3 ± 3.9Bab | 26.0 ± 4.9Bab | 10.7 ± 3.5Ba | 5.0 ± 0.2ABb | 0.09 ± 0.02Ab | 1.9 ± 0.7Bb | 17.0 ± 7.5ABab | 72.4 ± 47.2Bb | 4.8 ± 2.84Bbc | |
| 30-40 | 1.7 ± 0.0Bb | 63.4 ± 3.7Bab | 25.8 ± 5.0Bab | 10.8 ± 3.4Ba | 5.1 ± 0.2Ab | 0.10 ± 0.03Ab | 1.6 ± 0.7ABb | 12.5 ± 3.2Ab | 40.7 ± 17.7ABc | 3.5 ± 1.6ABc | |
| PLL | 0-10 | 1.2 ± 0.3Aa | 49.0 ± 18.5Aa | 35.2 ± 14.0Ca | 15.8 ± 6.3Ba | 4.4 ± 0.2Ba | 0.15 ± 0.05Aa | 2.5 ± 0.9ABa | 21.5 ± 3.4Aa | 217.6 ± 54.6Ca | 10.2 ± 2.7Ba |
| 10-20 | 1.4 ± 0.2Aa | 45.2 ± 19.3Aa | 43.3 ± 18.0Ca | 11.6 ± 8.3Ba | 4.5 ± 0.3Bab | 0.13 ± 0.06Aa | 2.0 ± 0.4BCab | 20.1 ±6.5Aab | 103.6 ± 33.3Bb | 5.5 ± 2.1Bb | |
| 20-30 | 1.5 ± 0.2Aa | 43.0 ± 18.2Aa | 45.6 ± 19.4Ca | 11.5 ± 6.8Ba | 4.6 ± 0.3Bab | 0.11 ± 0.06Aa | 1.7 ± 0.8ABb | 16.6 ± 4.9ABb | 72.7 ± 26.5Bc | 4.5 ± 1.7Bbc | |
| 30-40 | 1.5 ± 0.1Aa | 41.2 ± 18.7Aa | 48.8 ± 19.7Aa | 10.1 ± 7.9Ba | 4.7 ± 0.3Ab | 0.13 ± 0.08Aa | 2.0 ± 1.5Bab | 17.1 ± 5.6Bb | 46.8 ± 22.4ABc | 3.1 ± 2.1Ac | |
| LAG | 0-10 | 1.3 ± 0.1Aa | 26.1 ± 7.4Ca | 56.5 ± 8.4Da | 17.4 ± 4.2Ba | 4.4 ± 0.3Ca | 0.25 ± 0.05Ba | 2.8 ± 0.9Ba | 16.5 ± 2.9Aa | 151.4 ± 55.8Ba | 8.6 ± 3.7Ba |
| 10-20 | 1.4 ± 0.0Ab | 20.1 ± 3.6Cb | 61.2 ± 4.8Dab | 18.8 ± 4.8Ca | 4.6 ± 0.2Ca | 0.20 ± 0.04Bb | 2.5 ± 0.9Cab | 13.2 ± 1.2Bb | 84.7 ± 27.3ABb | 6.5 ± 2.1Bb | |
| 20-30 | 1.5 ± 0.0Ab | 21.8 ± 5.2Cab | 62.9 ± 5.0Ab | 15.3 ± 4.3Cab | 4.6 ± 0.2Ca | 0.19 ± 0.04Bb | 2.0 ± 1.0Bb | 11.9 ± 1.6Bb | 67.4 ± 25.0ABb | 5.6 ± 1.8Bb | |
| 30-40 | 1.5 ± 0.0Ab | 23.9 ± 6.3Cab | 62.5 ± 7.3Cb | 13.6 ± 4.5Bb | 4.6 ± 0.3Ca | 0.20 ± 0.05Bb | 1.9 ± 0.9ABb | 12.0 ± 1.7Ab | 63.3 ± 31.3Bb | 5.2 ± 2.4Bb |
表3 不同植被恢复阶段土壤的理化性质(平均值±标准偏差)
Table 3 Soil physicochemical properties at different stages during vegetation restoration (mean ± SD)
| 恢复 阶段 Restoration stage | 土层 深度 Soil layer (cm) | 容重 Bulk density (g·cm-3) | 0.05-2 mm 砂粒百分含量 Soil sand (0.05-2 mm) percentage (%) | 0.002-0.05 mm粉粒百分含量 Soil silt (0.002- 0.05 mm) percentage (%) | <0.002 mm 黏粒百分含量 Soil clay (<0.002 mm) percentage (%) | pH | TP (g·kg-1) | AP (mg·kg-1) | C/N | C/P | N/P |
|---|---|---|---|---|---|---|---|---|---|---|---|
| LVR | 0-10 | 1.4 ± 0.2Aa | 43.6 ± 4.8Aa | 44.7 ± 6.4Aa | 11.7 ± 4.6Aa | 4.5 ± 0.2Aa | 0.17 ± 0.16Aa | 2.1 ± 0.7Aa | 22.4 ± 12.5Aa | 109.1 ± 44.7Aa | 5.3 ± 2.4Aa |
| 10-20 | 1.5 ± 0.1Aab | 39.5 ± 4.1Aa | 57.0 ± 5.2Ab | 3.5 ± 1.1Ab | 4.7 ± 0.2Ab | 0.12 ± 0.03Aab | 1.4 ± 0.6Ab | 20.0 ± 9.3Aab | 59.5 ± 51.4Ab | 3.3 ± 3.0Ab | |
| 20-30 | 1.5 ± 0.1Aab | 42.0 ± 5.6Aa | 56.5 ± 6.2Ab | 2.7 ± 0.9Ab | 4.8 ± 0.2Ab | 0.11 ± 0.04Ab | 1.3 ± 0.5Ab | 17.7 ± 9.5Aab | 42.9 ± 40.9Ab | 2.5 ± 2.2Ab | |
| 30-40 | 1.5 ± 0.1Ab | 43.1 ± 5.6Aa | 54.4 ± 6.1ACb | 2.5 ± 0.6Ab | 5.0 ± 0.2Ac | 0.11 ± 0.04Ab | 1.2 ± 0.5Ab | 13.7 ± 7.7ABb | 30.5 ± 37.4Ab | 2.5 ± 2.5Ab | |
| LCQ | 0-10 | 1.4 ± 0.1Ab | 67.9 ± 2.1Ba | 21.6 ± 2.5Ba | 10.6 ± 2.4Aa | 4.8 ± 0.2Ba | 0.12 ± 0.03Aa | 2.6 ± 1.1ABa | 17.7 ± 6.8Aa | 164.4 ± 45.5Ba | 10.0 ± 3.1Ba |
| 10-20 | 1.6 ± 0.1Ab | 60.1 ± 11.1Bb | 27.2 ± 9.1Bb | 12.9 ± 5.0Ba | 4.9 ± 0.2ABab | 0.10 ± 0.03Ab | 1.9 ± 0.6Bab | 16.3 ± 4.6ABab | 90.2 ± 29.9Bb | 6.0 ± 2.8Bb | |
| 20-30 | 1.6 ± 0.1Ab | 63.3 ± 3.9Bab | 26.0 ± 4.9Bab | 10.7 ± 3.5Ba | 5.0 ± 0.2ABb | 0.09 ± 0.02Ab | 1.9 ± 0.7Bb | 17.0 ± 7.5ABab | 72.4 ± 47.2Bb | 4.8 ± 2.84Bbc | |
| 30-40 | 1.7 ± 0.0Bb | 63.4 ± 3.7Bab | 25.8 ± 5.0Bab | 10.8 ± 3.4Ba | 5.1 ± 0.2Ab | 0.10 ± 0.03Ab | 1.6 ± 0.7ABb | 12.5 ± 3.2Ab | 40.7 ± 17.7ABc | 3.5 ± 1.6ABc | |
| PLL | 0-10 | 1.2 ± 0.3Aa | 49.0 ± 18.5Aa | 35.2 ± 14.0Ca | 15.8 ± 6.3Ba | 4.4 ± 0.2Ba | 0.15 ± 0.05Aa | 2.5 ± 0.9ABa | 21.5 ± 3.4Aa | 217.6 ± 54.6Ca | 10.2 ± 2.7Ba |
| 10-20 | 1.4 ± 0.2Aa | 45.2 ± 19.3Aa | 43.3 ± 18.0Ca | 11.6 ± 8.3Ba | 4.5 ± 0.3Bab | 0.13 ± 0.06Aa | 2.0 ± 0.4BCab | 20.1 ±6.5Aab | 103.6 ± 33.3Bb | 5.5 ± 2.1Bb | |
| 20-30 | 1.5 ± 0.2Aa | 43.0 ± 18.2Aa | 45.6 ± 19.4Ca | 11.5 ± 6.8Ba | 4.6 ± 0.3Bab | 0.11 ± 0.06Aa | 1.7 ± 0.8ABb | 16.6 ± 4.9ABb | 72.7 ± 26.5Bc | 4.5 ± 1.7Bbc | |
| 30-40 | 1.5 ± 0.1Aa | 41.2 ± 18.7Aa | 48.8 ± 19.7Aa | 10.1 ± 7.9Ba | 4.7 ± 0.3Ab | 0.13 ± 0.08Aa | 2.0 ± 1.5Bab | 17.1 ± 5.6Bb | 46.8 ± 22.4ABc | 3.1 ± 2.1Ac | |
| LAG | 0-10 | 1.3 ± 0.1Aa | 26.1 ± 7.4Ca | 56.5 ± 8.4Da | 17.4 ± 4.2Ba | 4.4 ± 0.3Ca | 0.25 ± 0.05Ba | 2.8 ± 0.9Ba | 16.5 ± 2.9Aa | 151.4 ± 55.8Ba | 8.6 ± 3.7Ba |
| 10-20 | 1.4 ± 0.0Ab | 20.1 ± 3.6Cb | 61.2 ± 4.8Dab | 18.8 ± 4.8Ca | 4.6 ± 0.2Ca | 0.20 ± 0.04Bb | 2.5 ± 0.9Cab | 13.2 ± 1.2Bb | 84.7 ± 27.3ABb | 6.5 ± 2.1Bb | |
| 20-30 | 1.5 ± 0.0Ab | 21.8 ± 5.2Cab | 62.9 ± 5.0Ab | 15.3 ± 4.3Cab | 4.6 ± 0.2Ca | 0.19 ± 0.04Bb | 2.0 ± 1.0Bb | 11.9 ± 1.6Bb | 67.4 ± 25.0ABb | 5.6 ± 1.8Bb | |
| 30-40 | 1.5 ± 0.0Ab | 23.9 ± 6.3Cab | 62.5 ± 7.3Cb | 13.6 ± 4.5Bb | 4.6 ± 0.3Ca | 0.20 ± 0.05Bb | 1.9 ± 0.9ABb | 12.0 ± 1.7Ab | 63.3 ± 31.3Bb | 5.2 ± 2.4Bb |
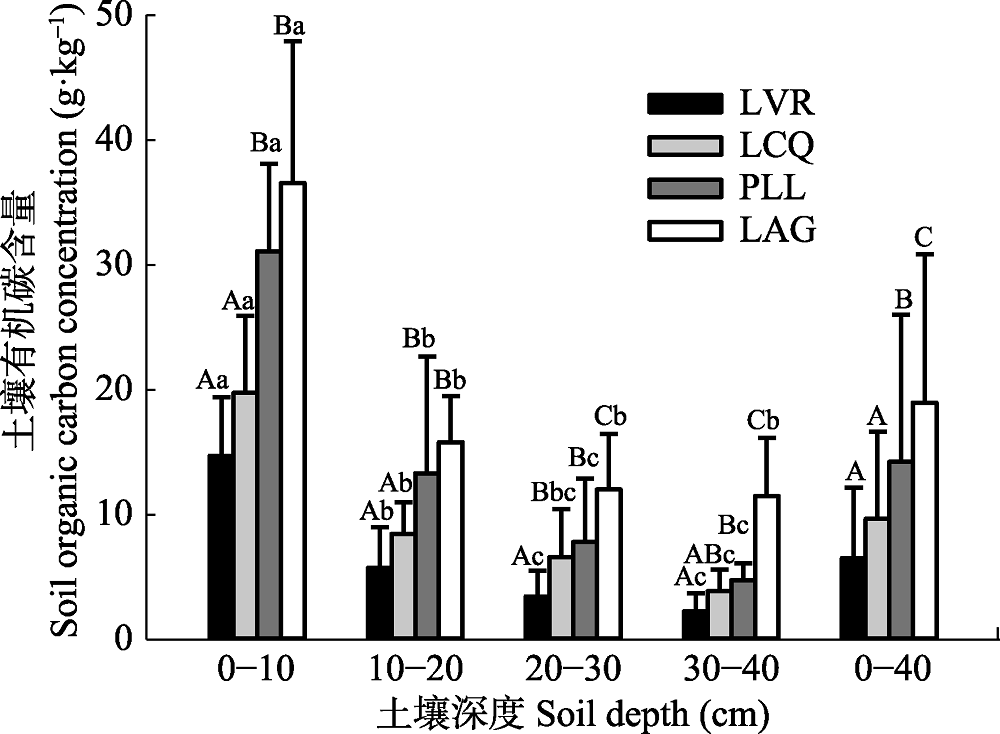
图1 不同植被恢复阶段的土壤有机碳含量(平均值±标准偏差)。LAG, 柯-红淡比-青冈常绿阔叶林; LCQ, 檵木-杉木-白栎灌木林; LVR, 檵木-南烛-杜鹃灌草丛; PLL, 马尾松-柯-檵木针阔混交林。不同大写字母表示同一土层不同植被恢复阶段之间差异显著(p < 0.05), 不同小写字母表示同一植被恢复阶段不同土层之间差异显著(p < 0.05)。
Fig. 1 Soil organic carbon concentration at different stages during vegetation restoration (mean ± SD). LVR, Loropetalum chinense-Vaccinium bracteatum-Rhododendron simsii scrub- grass-land; LCQ, Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery; PLL, Pinus massoniana- Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest; LAG, Lithocarpus glaber-Cleyera japonica- Cyclobalanopsis glauca evergreen broad-leaved forest. Different capital letters represent significant differences among different vegetation restoration stages in the same soil layer (p < 0.05), and different lower letters indicate significant differences among different soil layers at the same vegetation restoration stage (p < 0.05).
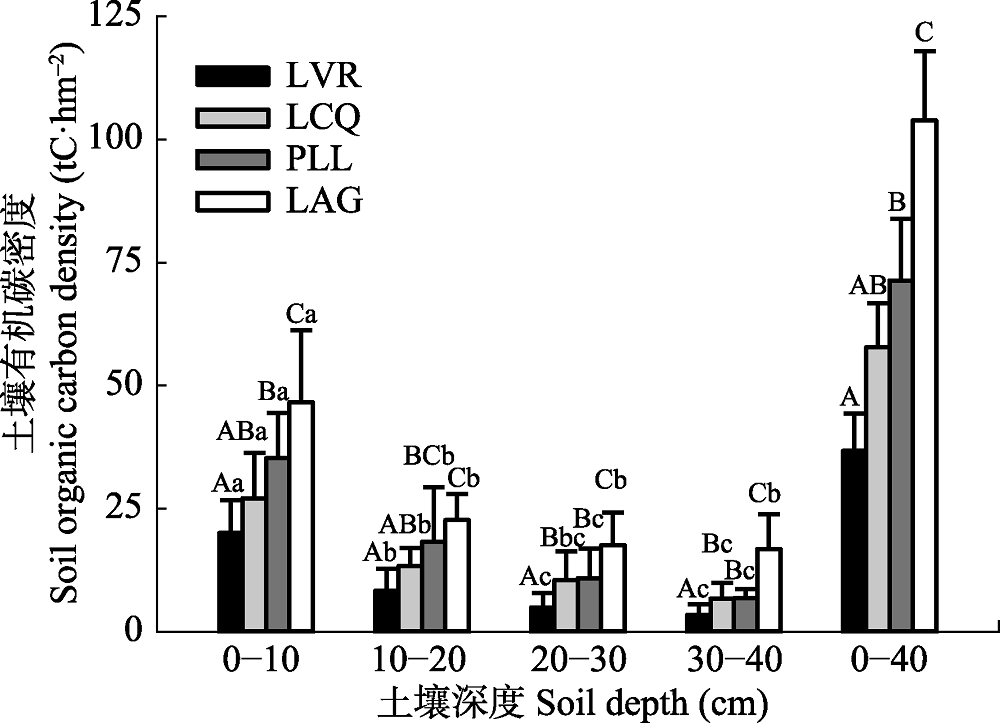
图2 不同植被恢复阶段的土壤有机碳密度(平均值±标准偏差)。图注同图1。
Fig. 2 Soil organic carbon density at different stages during vegetation restoration (mean ± SD). See Fig. 1 for notes.
| 项目 Item | 植物多样性指数Species diversity index | 群落总 生物量 Community total biomass | 地上部分 生物量 Aboveground biomass | 根系生 物量 Root biomass | 凋落物层 现存量 Existing biomass in litter layer | 凋落物层C含量 C concentration in litter layer | 凋落物层N含量 N concentration in litter layer | 凋落物层P含量 P concentration in litter layer | 凋落物层C/N比 C/N ratio in litter layer | 凋落物层C/P比 C/P ratio in litter layer | 凋落物层N/P比 N/P ratio in litter layer |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CSOC | 0.293* | 0.479** | 0.473** | 0.478** | 0.456** | 0.162 | 0.459** | 0.378** | -0.240 | -0.279* | -0.070 |
| DSOC | 0.326* | 0.476** | 0.468** | 0.495** | 0.470** | 0.102 | 0.496** | 0.424** | -0.318* | -0.348* | -0.083 |
表4 土壤有机碳含量(CSOC)、密度(DSOC)与植被因子的Pearson相关系数
Table 4 Pearson’s correlation coefficient between soil organic carbon concentration (CSOC), density (DSOC) and vegetation factors
| 项目 Item | 植物多样性指数Species diversity index | 群落总 生物量 Community total biomass | 地上部分 生物量 Aboveground biomass | 根系生 物量 Root biomass | 凋落物层 现存量 Existing biomass in litter layer | 凋落物层C含量 C concentration in litter layer | 凋落物层N含量 N concentration in litter layer | 凋落物层P含量 P concentration in litter layer | 凋落物层C/N比 C/N ratio in litter layer | 凋落物层C/P比 C/P ratio in litter layer | 凋落物层N/P比 N/P ratio in litter layer |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CSOC | 0.293* | 0.479** | 0.473** | 0.478** | 0.456** | 0.162 | 0.459** | 0.378** | -0.240 | -0.279* | -0.070 |
| DSOC | 0.326* | 0.476** | 0.468** | 0.495** | 0.470** | 0.102 | 0.496** | 0.424** | -0.318* | -0.348* | -0.083 |
| 项目 Item | 容重 Bulk density | 0.05-2 mm 砂粒百分含量 Soil sand percentage (0.05-2 mm) | 0.002-0.05 mm 粉粒百分含量 Soil silt percentage (0.002-0.05 mm) | <0.002 mm 黏粒百分含量 Soil clay percentage (<0.002 mm) | pH | TP | AP | C/N | C/P | N/P |
|---|---|---|---|---|---|---|---|---|---|---|
| CSOC | -0.678** | -0.246 | 0.027 | 0.637** | -0.666** | 0.568** | 0.727** | 0.277* | 0.792** | 0.780** |
| DSOC | -0.551** | -0.209 | -0.007 | 0.623** | -0.623** | 0.564** | 0.752** | 0.2430 | 0.796** | 0.811** |
表5 土壤有机碳含量(CSOC)、密度(DSOC)与土壤因子的Pearson相关系数
Table 5 Pearson’s correlation coefficient between soil organic carbon concentration (CSOC), density (DSOC) and soil factors
| 项目 Item | 容重 Bulk density | 0.05-2 mm 砂粒百分含量 Soil sand percentage (0.05-2 mm) | 0.002-0.05 mm 粉粒百分含量 Soil silt percentage (0.002-0.05 mm) | <0.002 mm 黏粒百分含量 Soil clay percentage (<0.002 mm) | pH | TP | AP | C/N | C/P | N/P |
|---|---|---|---|---|---|---|---|---|---|---|
| CSOC | -0.678** | -0.246 | 0.027 | 0.637** | -0.666** | 0.568** | 0.727** | 0.277* | 0.792** | 0.780** |
| DSOC | -0.551** | -0.209 | -0.007 | 0.623** | -0.623** | 0.564** | 0.752** | 0.2430 | 0.796** | 0.811** |
| 因子 Factor | 主成分 Component | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| 植物多样性指数 Species diversity index | 0.687 | -0.504 | -0.200 | -0.020 | -0.259 |
| 群落总生物量 Community total biomass | 0.877 | 0.247 | -0.056 | 0.301 | 0.169 |
| 地上部分生物量 Aboveground biomass | 0.862 | 0.279 | -0.054 | 0.311 | 0.162 |
| 根系部分生物量 Root biomass | 0.913 | 0.132 | -0.13 | 0.221 | 0.221 |
| 凋落物层现存量 Existing biomass in litter layer | 0.841 | -0.247 | 0.112 | 0.180 | 0.079 |
| 凋落物层C含量 C concentration in litter layer | 0.281 | 0.562 | 0.149 | 0.708 | 0.001 |
| 凋落物层N含量 N concentration in litter layer | 0.786 | -0.123 | -0.232 | -0.011 | 0.218 |
| 凋落物层P含量 P concentration in litter layer | 0.810 | -0.360 | -0.080 | 0.199 | -0.263 |
| 凋落物层C/N比 C/N ratio in litter layer | -0.522 | 0.625 | 0.241 | 0.501 | -0.106 |
| 凋落物层C/P比 C/P ratio in litter layer | -0.622 | 0.745 | 0.06 | 0.12 | 0.112 |
| 凋落物层N/P比 N/P ratio in litter layer | -0.204 | 0.439 | -0.3 | -0.533 | 0.577 |
| 土壤容重 Soil bulk density | -0.384 | -0.614 | -0.367 | 0.237 | 0.157 |
| 0.05-2 mm砂粒百分含量 Soil sand percentage (0.05-2.00 mm) | -0.531 | -0.57 | 0.549 | 0.187 | 0.078 |
| 0.002-0.05 mm粉粒百分含量 Soil silt percentage (0.002-0.05 mm) | 0.276 | 0.615 | -0.65 | -0.122 | -0.098 |
| <0.002 mm黏粒百分含量 Soil clay percentage (<0.002 mm) | 0.914 | -0.023 | 0.174 | -0.21 | 0.043 |
| 土壤pH值 Soil pH value | -0.575 | -0.702 | -0.204 | 0.159 | 0.195 |
| 土壤全磷含量 Soil total phosphorus concentration | 0.631 | 0.342 | -0.248 | -0.457 | -0.291 |
| 土壤速效磷含量 Soil available phosphorus concentration | 0.656 | -0.095 | 0.482 | -0.204 | -0.187 |
| 土壤碳氮比 Soil C/N ratio | -0.244 | 0.446 | 0.616 | -0.205 | -0.273 |
| 土壤碳磷比 Soil C/P ratio | 0.350 | 0.066 | 0.861 | -0.155 | 0.191 |
| 土壤氮磷比 Soil N/P ratio | 0.499 | -0.120 | 0.720 | -0.172 | 0.296 |
| 贡献率 Contribution rate | 40.533 | 18.984 | 15.113 | 8.953 | 4.995 |
| 累积贡献率 Accumulative contribution rate | 40.533 | 59.517 | 74.630 | 83.583 | 88.578 |
表6 植被因子和土壤因子主成分载荷矩阵、特征值及贡献率
Table 6 The principle component loading matrix, eigenvalue, contribution rate for vegetation and soil factors
| 因子 Factor | 主成分 Component | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| 植物多样性指数 Species diversity index | 0.687 | -0.504 | -0.200 | -0.020 | -0.259 |
| 群落总生物量 Community total biomass | 0.877 | 0.247 | -0.056 | 0.301 | 0.169 |
| 地上部分生物量 Aboveground biomass | 0.862 | 0.279 | -0.054 | 0.311 | 0.162 |
| 根系部分生物量 Root biomass | 0.913 | 0.132 | -0.13 | 0.221 | 0.221 |
| 凋落物层现存量 Existing biomass in litter layer | 0.841 | -0.247 | 0.112 | 0.180 | 0.079 |
| 凋落物层C含量 C concentration in litter layer | 0.281 | 0.562 | 0.149 | 0.708 | 0.001 |
| 凋落物层N含量 N concentration in litter layer | 0.786 | -0.123 | -0.232 | -0.011 | 0.218 |
| 凋落物层P含量 P concentration in litter layer | 0.810 | -0.360 | -0.080 | 0.199 | -0.263 |
| 凋落物层C/N比 C/N ratio in litter layer | -0.522 | 0.625 | 0.241 | 0.501 | -0.106 |
| 凋落物层C/P比 C/P ratio in litter layer | -0.622 | 0.745 | 0.06 | 0.12 | 0.112 |
| 凋落物层N/P比 N/P ratio in litter layer | -0.204 | 0.439 | -0.3 | -0.533 | 0.577 |
| 土壤容重 Soil bulk density | -0.384 | -0.614 | -0.367 | 0.237 | 0.157 |
| 0.05-2 mm砂粒百分含量 Soil sand percentage (0.05-2.00 mm) | -0.531 | -0.57 | 0.549 | 0.187 | 0.078 |
| 0.002-0.05 mm粉粒百分含量 Soil silt percentage (0.002-0.05 mm) | 0.276 | 0.615 | -0.65 | -0.122 | -0.098 |
| <0.002 mm黏粒百分含量 Soil clay percentage (<0.002 mm) | 0.914 | -0.023 | 0.174 | -0.21 | 0.043 |
| 土壤pH值 Soil pH value | -0.575 | -0.702 | -0.204 | 0.159 | 0.195 |
| 土壤全磷含量 Soil total phosphorus concentration | 0.631 | 0.342 | -0.248 | -0.457 | -0.291 |
| 土壤速效磷含量 Soil available phosphorus concentration | 0.656 | -0.095 | 0.482 | -0.204 | -0.187 |
| 土壤碳氮比 Soil C/N ratio | -0.244 | 0.446 | 0.616 | -0.205 | -0.273 |
| 土壤碳磷比 Soil C/P ratio | 0.350 | 0.066 | 0.861 | -0.155 | 0.191 |
| 土壤氮磷比 Soil N/P ratio | 0.499 | -0.120 | 0.720 | -0.172 | 0.296 |
| 贡献率 Contribution rate | 40.533 | 18.984 | 15.113 | 8.953 | 4.995 |
| 累积贡献率 Accumulative contribution rate | 40.533 | 59.517 | 74.630 | 83.583 | 88.578 |
| 因子 Factor | 模型 Model | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| 土壤碳磷比 Soil C/P ratio | 0.141 | 0.111 | 0.101 |
| 土壤pH值 Soil pH value | -17.073 | -19.492 | |
| 凋落物层C/P比 C/P ratio in litter layer | -0.009 | ||
| 常数项 Constant term | -0.285 | 82.592 | 105.516 |
| 多元相关系数 Multiple correlation coefficient (R) | 0.792 | 0.874 | 0.912 |
| 调整判定系数 Adjust R2 | 0.620 | 0.755 | 0.820 |
| F | 84.203 | 79.603 | 78.584 |
| p | 0.000 | 0.000 | 0.000 |
表7 土壤有机碳含量影响因子的逐步回归分析
Table 7 The stepwise regression analysis for main influencing factors of soil organic carbon concentration
| 因子 Factor | 模型 Model | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| 土壤碳磷比 Soil C/P ratio | 0.141 | 0.111 | 0.101 |
| 土壤pH值 Soil pH value | -17.073 | -19.492 | |
| 凋落物层C/P比 C/P ratio in litter layer | -0.009 | ||
| 常数项 Constant term | -0.285 | 82.592 | 105.516 |
| 多元相关系数 Multiple correlation coefficient (R) | 0.792 | 0.874 | 0.912 |
| 调整判定系数 Adjust R2 | 0.620 | 0.755 | 0.820 |
| F | 84.203 | 79.603 | 78.584 |
| p | 0.000 | 0.000 | 0.000 |
| 因子 Factor | 模型 Model | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| 土壤C/P比 Soil C/P ratio | 0.172 | 0.141 | 0.127 | 0.124 |
| <0.002 mm黏粒百分含量 Soil clay percentage (< 0.002 mm) | 0.763 | 0.565 | 0.107 | |
| 土壤pH值 Soil pH value | -12.315 | -20.204 | ||
| 凋落物层C/P比 C/P ratio in litter layer | -0.012 | |||
| 常数项 Constant term | 1.416 | -4.094 | 57.112 | 114.025 |
| 多元相关系数 Multiple correlation coefficient (R) | 0.796 | 0.865 | 0.889 | 0.916 |
| 调整判定系数 Adjust (R2) | 0.626 | 0.739 | 0.776 | 0.825 |
| F | 86.325 | 73.058 | 60.022 | 61.066 |
| p | 0.000 | 0.000 | 0.000 | 0.000 |
表8 土壤有机碳密度影响因子的逐步回归分析
Table 8 The stepwise regression analysis for main influencing factors of soil organic carbon density
| 因子 Factor | 模型 Model | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| 土壤C/P比 Soil C/P ratio | 0.172 | 0.141 | 0.127 | 0.124 |
| <0.002 mm黏粒百分含量 Soil clay percentage (< 0.002 mm) | 0.763 | 0.565 | 0.107 | |
| 土壤pH值 Soil pH value | -12.315 | -20.204 | ||
| 凋落物层C/P比 C/P ratio in litter layer | -0.012 | |||
| 常数项 Constant term | 1.416 | -4.094 | 57.112 | 114.025 |
| 多元相关系数 Multiple correlation coefficient (R) | 0.796 | 0.865 | 0.889 | 0.916 |
| 调整判定系数 Adjust (R2) | 0.626 | 0.739 | 0.776 | 0.825 |
| F | 86.325 | 73.058 | 60.022 | 61.066 |
| p | 0.000 | 0.000 | 0.000 | 0.000 |
| [1] |
Aerts R ( 1997). Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos, 79, 439-449.
DOI URL |
| [2] |
Berger TW, Neubauer C, Glatzel G ( 2002). Factors controlling soil carbon and nitrogen stores in pure stands of Norway spruce (Picea abies) and mixed species stands in Austria. Forest Ecology and Management, 159, 3-14.
DOI URL |
| [3] | Bi JD, Li YL, Ning ZY, Zhao XY ( 2016). Carbon mineralization and decomposition of litters from dominant plants in the Horqin Sandy Land: Effect of litter quality. Journal of Desert Research, 36, 85-92. |
| [ 毕京东, 李玉霖, 宁志英, 赵学勇 ( 2016). 科尔沁沙地优势植物叶凋落物分解及碳矿化——凋落物质量的影响. 中国沙漠, 36, 85-92.] | |
| [4] |
Cao J, Yan WD, Xiang WH, Chen XY, Lei PF ( 2015). Stoichiometry characterization of soil C, N and P of Chinese fir plantations at three different ages in Huitong, Hunan Province, China. Scientia Silvae Sinicae, 51(7), 1-8.
DOI URL |
|
[ 曹娟, 闫文德, 项文化, 谌小勇, 雷丕锋 ( 2015). 湖南会同3个林龄杉木人工林土壤碳、氮、磷化学计量特征. 林业科学, 51(7), 1-8.]
DOI URL |
|
| [5] |
Castro H, Fortunel C, Freitas H ( 2010). Effects of land abandonment on plant litter decomposition in a Montado system: Relation to litter chemistry and community functional parameters. Plant and Soil, 333, 181-190.
DOI URL |
| [6] |
Chen J, Yang N ( 2016). Effect of re-vegetation on active organic carbon pool of purple soils on sloping-land in Hengyang, Hunan Province, China. Journal of Tropical and Subtropical Botany, 24, 568-576.
DOI URL |
|
[ 陈璟, 杨宁 ( 2016). 衡阳紫色土丘陵坡地植被恢复对土壤活性有机碳库的影响. 热带亚热带植物学报, 24, 568-576.]
DOI URL |
|
| [7] | Chen LZ, Xie BY, Xiao WF, Huang ZL ( 2007). Organic carbon storage in soil under the major forest vegetation types in the Three Gorges Reservior area. Resources and Environment in the Yangtze Basin, 16, 640-643. |
| [ 陈亮中, 谢宝元, 肖文发, 黄志霖 ( 2007). 三峡库区主要森林植被类型土壤有机碳贮量研究. 长江流域资源与环境, 16, 640-643.] | |
| [8] |
Chen S, Huang Y, Zou J, Shi Y ( 2013). Mean residence time of global topsoil organic carbon depends on temperature, precipitation and soil nitrogen. Global and Planetary Change, 100, 99-108.
DOI URL |
| [9] | Dai QH, Liu GB, Xue S, Yu N, Zhang C, Lan X ( 2008). Effect of different vegetation restoration on soil carbon and carbon management index in eroded Hilly Loess Plateau. Research of Soil and Water Conservation, 15(3), 61-64. |
| [ 戴全厚, 刘国彬, 薛萐, 于娜, 张超, 兰雪 ( 2008). 不同植被恢复模式对黄土丘陵区土壤碳库及其管理指数的影响. 水土保持研究, 15(3), 61-64.] | |
| [10] |
Deng L, Wang KB, Chen ML, Shangguan ZP, Sweeney S ( 2013). Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China. Catena, 110, 1-7.
DOI URL |
| [11] | Ding FJ, Pan ZS, Zhou FJ, Wu P ( 2012). Organic carbon contents and vertical distribution characteristics of the soil in the three forest types of the karst regions in central Guizhou Province. Journal of Soil and Water Conservation, 26(1), 161-164. |
| [ 丁访军, 潘忠松, 周凤娇, 吴鹏 ( 2012). 黔中喀斯特地区3种林型土壤有机碳含量及垂直分布特征. 水土保持学报, 26(1), 161-164.] | |
| [12] |
Du H, Zeng FP, Song TQ, Wen YG, Li CG, Peng WX, Zhang H, Zeng ZX ( 2016). Spatial pattern of soil organic carbon of the main forest soils and its influencing factors in Guangxi, China. Chinese Journal of Plant Ecology, 40, 282-291.
DOI URL |
|
[ 杜虎, 曾馥平, 宋同清, 温远光, 李春干, 彭晚霞, 张浩, 曾昭霞 ( 2016). 广西主要森林土壤有机碳空间分布及其影响因素. 植物生态学报, 40, 282-291.]
DOI URL |
|
| [13] |
Fang J, Chen A, Peng C, Zhao S, Ci L ( 2001). Changes in forest biomass carbon storage in China between 1949 and 1998. Science, 292, 2320-2322.
DOI URL PMID |
| [14] |
Fisk M, Santangelo S, Minick K ( 2015). Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests. Soil Biology & Biochemistry, 81, 212-218.
DOI URL |
| [15] | Fu DL, Liu MY, Liu L, Zhang K, Zuo JX ( 2014). Organic carbon density and storage in different soils on the Loess Plateau. Arid Zone Research, 1, 44-50. |
| [ 付东磊, 刘梦云, 刘林, 张琨, 左进香 ( 2014). 黄土高原不同土壤类型有机碳密度与储量特征. 干旱区研究, 31, 44-50.] | |
| [16] |
Gong W, Hu XT, Wang JY, Gong YB, Ran H ( 2008). Soil carbon pool and fertility under natural evergreen broad-leaved forest and its artificial regeneration forests in Southern Sichuan Province. Acta Ecologica Sinica, 28, 2536-2545.
DOI URL |
|
[ 龚伟, 胡庭兴, 王景燕, 宫渊波, 冉华 ( 2008). 川南天然常绿阔叶林人工更新后土壤碳库与肥力的变化. 生态学报, 28, 2536-2545.]
DOI URL |
|
| [17] |
Grüneberg E, Schöning I, Hessenmöller D, Schulze ED, Weisser WW ( 2013). Organic layer and clay content control soil organic carbon stocks in density fractions of differently managed German beech forests. Forest Ecology and Management, 303, 1-10.
DOI URL |
| [18] | Gu X, Fang X, Xiang WH, Li SL, Sun WJ ( 2013). Soil organic carbon and mineralizable organic carbon in four subtropical forests in hilly region of central Hunan Province, China. Chinese Journal of Ecology, 32, 2687-2694. |
| [ 辜翔, 方晰, 项文化, 李胜蓝, 孙伟军 ( 2013). 湘中丘陵区4种森林类型土壤有机碳和可矿化有机碳的比较. 生态学杂志, 32, 2687-2649.] | |
| [19] |
Guo SL, Ma YH, Che SG, Sun WY ( 2009). Effects of artificial and natural vegetations on litter production and soil organic carbon change in loess hilly areas. Scientia Silvae Sinicae, 45(10), 14-48.
DOI |
|
[ 郭胜利, 马玉红, 车升国, 孙文义 ( 2009). 黄土区人工与天然植被对凋落物量和土壤有机碳变化的影响. 林业科学, 45(10), 14-18.]
DOI |
|
| [20] |
Harrison-Kirk T, Beare MH, Meenken ED, Condron LM ( 2013). Soil organic matter and texture affect responses to dry/wet cycles: Effects on carbon dioxide and nitrous oxide emissions. Soil Biology & Biochemistry, 57, 43-55.
DOI URL |
| [21] |
Herbert DA, Williams M, Rastetter EB ( 2003). Amodel analysis of N and P limitation on carbon accumulation in Amazonian secondary forest after alternate land-use abandonment. Biogeochemistry, 65, 121-150.
DOI URL |
| [22] | Huang ZS, Yu LF, Fu YH ( 2012). Characteristics of soil mineralizable carbon pool in natural restoration process of karst forest vegetation. Chinese Journal of Applied Ecology, 23, 2165-2170. |
| [ 黄宗胜, 喻理飞, 符裕红 ( 2012). 喀斯特森林植被自然恢复过程中土壤可矿化碳库特征. 应用生态学报, 23, 2165-2170.] | |
| [23] |
Jha P, De A, Lakaria BL, Biswas AK, Singh M, Reddy KS, Rao AS ( 2012). Soil carbon pools, mineralization and fluxes associated with land use change in vertisols of Central India. Natinal Academy Science Letters, 35, 475-483.
DOI URL |
| [24] |
Jia GM, Cao J, Wang C, Wang G ( 2005). Microbial biomass and nutrients in soils at the different stages of secondary forest succession in Ziwulin, northwest China. Forest Ecology and Management, 217, 117-125.
DOI URL |
| [25] |
Kunkel ML, Flores AN, Smith TJ, Mcnamara JP, Benner SG ( 2011). A simplified approach for estimating soil carbon and nitrogen stocks in semi-arid complex terrain. Geoderma, 165, 1-11.
DOI URL |
| [26] |
Lal R ( 1999). Soil management and restoration for C sequestration to mitigate the accelerated greenhouse effect. Progress in Environmental Science, 1, 307-326.
DOI URL |
| [27] | Li B, Fang X, Li Y, Xiang WH, Tian DL, Chen XY, Yan WD, Deng DH ( 2015). Dynamic properties of soil organic carbon in Hunan’s forests. Acta Ecologica Sinica, 35, 4265-4278. |
| [ 李斌, 方晰, 李岩, 项文化, 田大伦, 谌小勇, 闫文德, 邓东华 ( 2015). 湖南省森林土壤有机碳密度及碳库储量动态. 生态学报, 35, 4265-4278.] | |
| [28] | Li SJ, Qiu LP, Zhang XC ( 2010). Mineralization of soil organic carbon and its relations with soil physical and chemical properties on the Loess Plateau. Acta Ecologica Sinica, 30, 1217-1226. |
| [ 李顺姬, 邱莉萍, 张兴昌 ( 2010). 黄土高原土壤有机碳矿化及其与土壤理化性质的关系. 生态学报, 30, 1217-1226.] | |
| [29] | Liang QP, Yu XX, Pang Z, Wang C, Lü XZ ( 2010). Study on soil organic carbon density of different forest types. Ecology and Environmental Sciences, 19, 889-893. |
| [ 梁启鹏, 余新晓, 庞卓, 王琛, 吕锡芝 ( 2010). 不同林分土壤有机碳密度研究. 生态环境学报, 19, 889-893.] | |
| [30] |
Liu C, Xiang WH, Lei PF, Deng XW, Tian DL, Fang X, Peng CH ( 2014). Standing fine root mass and production in four Chinese subtropical forests along a succession and species diversity gradient. Plant and Soil, 376, 445-459.
DOI URL |
| [31] | Liu JS, Yang JS, Yu JB, Wang JD ( 2003). Study on vertical distribution of soil organic carbon in wetlands Sanjiang Plain. Journal of Soil and Water Conservation, 17(3), 5-8. |
| [ 刘景双, 杨继松, 于君宝, 王金达 ( 2003). 三江平原沼泽湿地土壤有机碳垂直分布特征研究. 水土保持学报, 17(3), 5-8.] | |
| [32] | Liu SR, Wang H, Luan JW ( 2011). A review of research progress and future prospective of forest soil carbon stock and soil carbon process in China. Acta Ecologica Sinica, 31, 5437-5448. |
| [ 刘世荣, 王晖, 栾军伟 ( 2011). 中国森林土壤碳储量与土壤碳过程研究进展. 生态学报, 31, 5437-5448.] | |
| [33] |
Liu W, Chen S, Qin X, Baumann F, Scholten T, Zhou Z, Sun W, Zhang T, Ren J, Qin D ( 2012). Storage, patterns, and control of soil organic carbon and nitrogen in the northeastern margin of the Qinghai-Tibetan Plateau. Environmental Research Letters, 7, 35401-35412.
DOI URL |
| [34] | Liu WD, Su JR, Li SF, Zhang ZJ, Li ZW ( 2010). Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Acta Ecologica Sinica, 30, 6581-6590. |
| [ 刘万德, 苏建荣, 李帅锋, 张志钧, 李忠文 ( 2010). 云南普洱季风常绿阔叶林演替系列植物和土壤 C、N、P 化学计量特征. 生态学报, 30, 6581-6590.] | |
| [35] | Liu ZH, Chen LX, Wang LL ( 2009). Soil active carbon in soils under different stages of Korean pine and broadleaf forest. Chinese Journal of Soil Science, 40, 1088-1103. |
| [ 刘振花, 陈立新, 王琳琳 ( 2009). 红松阔叶混交林不同演替阶段土壤活性有机碳的研究. 土壤通报, 40, 1088-1103.] | |
| [36] |
McKane RB, Rastetter EB, Melillo JM, Shaver GR, Hopkinson CS, Fernandes DN ( 1995). Effects of global change on carbon storage in tropical forests of South America. Global Biogeochemical Cycle, 9, 329-350.
DOI URL |
| [37] |
Motavalli PP, Palm CA, Parton CA, Elliott ET, Frey SD ( 1995). Soil pH and organic C dynamics in tropical forest soils: Evidence from laboratory and simulation studies. Soil Biology & Biochemistry, 27, 1589-1599.
DOI URL |
| [38] |
Post WM, Kwon KC ( 2000). Soil carbon sequestration and land-use change: Processes and Potential. Global Change Biology, 6, 317-327.
DOI URL |
| [39] |
Ramesh T, Manjaiah KM, Mohopatra KP, Rajasekar K, Ngachan SV ( 2015). Assessment of soil organic carbon stocks and fractions under different agroforestry systems in subtropical hill agroecosystems of north-east India. Agroforestry Systems, 89, 677-690.
DOI URL |
| [40] |
Reichstein M, Bahn M, Ciais P, Frank D, Mahecha1 MD, Seneviratne SI, Zscheischler J, Beer C, Buchmann N, Frank DC, Papale D, Rammig A, Smith P, Thonicke K, van der Velde M, Vicca S, Walz A, Wattenbach M ( 2013). Climate extremes and the carbon cycle. Nature, 500, 287-295.
DOI URL PMID |
| [41] |
Sá JCDM, Lal R ( 2009). Stratification ratio of soil organic matter pools as an indicator of carbon sequestration in a tillage chronosequence on a Brazilian Oxisol. Soil and Tillage Research, 103, 46-56.
DOI URL |
| [42] | Sun BW, Yang XD, Zhang ZH, Ma WJ, Arshad A, Huang HX, Yan ER ( 2013). Relationships between soil carbon pool and vegetation carbon return through succession of evergreen broad-leaved forests in Tiantong region, Zhejiang Province, Eastern China. Chinese Journal of Plant Ecology, 37, 803-810. |
| [ 孙宝伟, 杨晓东, 张志浩, 马文济, Ali Arshad, 黄海侠, 阎恩荣 ( 2013). 浙江天童常绿阔叶林演替过程中土壤碳库与植被碳归还的关系. 植物生态学报, 37, 803-810.] | |
| [43] |
Sun WJ, Fang X, Xiang WH, Zhang SJ, Li SL ( 2013). Active pools of soil organic carbon in subtropical forests at different successional stages in Central Hunan, China. Acta Ecologica Sinica, 33, 7765-7773.
DOI URL |
|
[ 孙伟军, 方晰, 项文化, 张仕吉, 李胜蓝 ( 2013). 湘中丘陵区不同演替阶段森林土壤活性有机碳库特征. 生态学报, 33, 7765-7773.]
DOI URL |
|
| [44] |
Tang PH, Dang KL, Wang LH, Ma J ( 2016). Factors affecting soil organic carbon density in Betula albo-sinensis forests on the southern slope of the Qinling Mountains. Acta Ecologica Sinica, 36, 1030-1039.
DOI URL |
|
[ 唐朋辉, 党坤良, 王连贺, 马俊 ( 2016). 秦岭南坡红桦林土壤有机碳密度影响因素. 生态学报, 36, 1030-1039.]
DOI URL |
|
| [45] |
Wang H, Hall CAS ( 2004). Modeling the effects of Hurricane Hugo on spatial and temporal variation in primary productivity and soil carbon and nitrogen in the Luquillo Experimental Forest, Puerto Rico. Plant and Soil, 263, 69-84.
DOI URL |
| [46] | Wang SF, Wang XK, Ouyang ZY ( 2012). Factors controlling soil organic carbon and total nitrogen densities in the upstream watershed of Miyun Reservoir, north China. Environmental Science, 33, 946-951. |
| [ 王淑芳, 王效科, 欧阳志云 ( 2012). 密云水库上游流域土壤有机碳和全氮密度影响因素研究. 环境科学, 33, 946-951.] | |
| [47] |
Wang YF, Fu BJ, Lü YH, Chen LD ( 2011). Effects of vegetation restoration on soil organic carbon sequestration at multiple scales in semi-arid Loess Plateau, China. Catena, 85, 58-66.
DOI URL |
| [48] |
Weintraub MN, Schimel JP ( 2003). Interactions between carbon and nitrogen mineralization and soil organic matter chemistry in arctic tundra soils. Ecosystems, 6, 129-143.
DOI URL |
| [49] |
Wynn JG, Bird MI, Vellen L, Grand-Clement E, Carter J, Berry SL ( 2006). Continental-scale measurement of the soil organic carbon pool with climatic, edaphic, and biotic controls. Global Biogeochemical Cycles, 20, 1-12.
DOI URL |
| [50] |
Xiang WH, Zhou J, Ouyang S, Zhang SL, Lei PF, Li JX, Deng XW, Fang X, Forrester DI ( 2016). Species-specific and general allometric equations for estimating tree biomass components of subtropical forests in southern China. European Journal of Forest Research, 135, 1-17.
DOI URL |
| [51] |
Xue S, Liu GB, Pan YP, Dai QH, Zhang C, Yu N ( 2009). Evolution of soil labile organic matter and carbon management index in the artificial Robinia of loess hilly area. Scientia Agricultura Sinica, 42, 1458-1464.
DOI URL |
|
[ 薛萐, 刘国彬, 潘彦平, 戴全厚, 张超, 余娜 ( 2009). 黄土丘陵区人工刺槐林土壤活性有机碳与碳库管理指数演变. 中国农业科学, 42, 1458-1464.]
DOI URL |
|
| [52] |
You LH, Ye GF, Chen ZH, Luo MJ, Gao W, You SS ( 2014). Litter decomposition and initial nutrient content of major dominant tree species on coastal sandy areas. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 43, 585-591.
DOI URL |
|
[ 尤龙辉, 叶功富, 陈增鸿, 罗美娟, 高伟, 游水生 ( 2014). 滨海沙地主要优势树种的凋落物分解及其与初始养分含量的关系. 福建农林大学学报(自然科学版), 43, 585-591.]
DOI URL |
|
| [53] |
Zak DR, Holmes WE, Burton AJ, Pregitzer KS, Talhelm AF ( 2008). Simulated atmospheric NO -3 deposition increases soil organic matter by slowing decomposition . Ecological Applications, 18, 2016-2027.
DOI URL PMID |
| [54] |
Zhang X, Han SJ, Wang SQ, Gu Y, Yue LY, Feng Y, Geng SC, Chen ZJ ( 2016). Change of soil organic carbon fractions at different successional stages of Betula platyphylla forest in Changbai Mountains. Chinese Journal of Ecology, 35, 282-289.
DOI URL |
|
[ 张雪, 韩士杰, 王树起, 谷越, 岳琳艳, 冯月, 耿世聪, 陈志杰 ( 2016). 长白山白桦林不同演替阶段土壤有机碳组分的变化. 生态学杂志, 35, 282-289.]
DOI URL |
|
| [55] |
Zhao YG, Liu XF, Wang ZL, Zhao SW ( 2015). Soil organic carbon fractions and sequestration across a 150-yr secondary forest chronosequence on the Loess Plateau, China. Catena, 133, 303-308.
DOI URL |
| [56] |
Zhu LQ, Huang RZ, Duan HL, Jia L, Wang H, Huang SH, Yi ZQ, Zhang WF ( 2017). Effects of artificially restored forests on soil organic carbon and active organic carbon in eroded red soil. Acta Ecologica Sinica, 37, 249-257.
DOI URL |
|
[ 朱丽琴, 黄荣珍, 段洪浪, 贾龙, 王赫, 黄诗华, 易志强, 张文锋 ( 2017). 红壤侵蚀地不同人工恢复林对土壤总有机碳和活性有机碳的影响. 生态学报, 37, 249-257.]
DOI URL |
| [1] | 江康威 张青青 王亚菲 李宏 丁雨 杨永强 吐尔逊娜依·热依木. 放牧干扰下天山北坡中段植物功能群特征及其与土壤环境因子的关系[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 陈昭铨, 王明慧, 胡子涵, 郎学东, 何云琼, 刘万德. 云南普洱季风常绿阔叶林幼苗的群落构建机制[J]. 植物生态学报, 2024, 48(1): 68-79. |
| [3] | 陈雪萍, 赵学勇, 张晶, 王瑞雄, 卢建男. 基于地理探测器的科尔沁沙地植被NDVI时空变化特征及其驱动因素[J]. 植物生态学报, 2023, 47(8): 1082-1093. |
| [4] | 罗娜娜, 盛茂银, 王霖娇, 石庆龙, 何宇. 长期植被恢复对中国西南喀斯特石漠化土壤活性有机碳组分含量和酶活性的影响[J]. 植物生态学报, 2023, 47(6): 867-881. |
| [5] | 仲琦, 李曾燕, 马炜, 况雨潇, 邱岭军, 黎蕴洁, 涂利华. 氮添加和凋落物处理对华西雨屏区常绿阔叶林凋落叶分解的影响[J]. 植物生态学报, 2023, 47(5): 629-643. |
| [6] | 冯印成, 王云琦, 王玉杰, 王凯, 王松年, 王杰帅. 重庆缙云山针阔混交林水汽通量特征及其影响因子[J]. 植物生态学报, 2022, 46(8): 890-903. |
| [7] | 秦江环, 张春雨, 赵秀海. 基于温带针阔混交林植物-土壤反馈的Janzen- Connell假说检验[J]. 植物生态学报, 2022, 46(6): 624-631. |
| [8] | 郝建锋, 周润惠, 姚小兰, 喻静, 陈聪琳, 向琳, 王姚瑶, 苏天成, 齐锦秋. 二代野猪放牧对夹金山针阔混交林物种多样性与土壤理化性质的影响[J]. 植物生态学报, 2022, 46(2): 197-207. |
| [9] | 崔光帅, 罗天祥, 梁尔源, 张林. 干旱半干旱区灌丛对草本植物的促进作用研究进展[J]. 植物生态学报, 2022, 46(11): 1321-1333. |
| [10] | 薛金儒, 吕肖良. 黄土高原生态工程实施下基于日光诱导叶绿素荧光的植被恢复生产力效益评价[J]. 植物生态学报, 2022, 46(10): 1289-1304. |
| [11] | 朱华. 云南常绿阔叶林的植被地理研究[J]. 植物生态学报, 2021, 45(3): 224-241. |
| [12] | 李帅锋, 郎学东, 黄小波, 王艳红, 刘万德, 徐崇华, 苏建荣. 云南普洱30 hm2季风常绿阔叶林动态监测样地群丛数量分类[J]. 植物生态学报, 2020, 44(3): 236-247. |
| [13] | 邓梦达, 游健荣, 李家湘, 李雄, 杨静, 邓创发, 刘昂, 刘文剑, 丁聪, 谢勇, 周国辉, 喻勋林. 长株潭城市群生态绿心地区主要植被类型的群落特征[J]. 植物生态学报, 2020, 44(12): 1296-1304. |
| [14] | 王艳红, 李帅锋, 郎学东, 黄小波, 刘万德, 徐崇华, 苏建荣. 地形异质性对云南普洱季风常绿阔叶林物种多样性的影响[J]. 植物生态学报, 2020, 44(10): 1015-1027. |
| [15] | 莫丹, 王振孟, 左有璐, 向双. 亚热带常绿阔叶林木本植物幼树阶段抽枝展叶的权衡关系[J]. 植物生态学报, 2020, 44(10): 995-1006. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19