
植物生态学报 ›› 2007, Vol. 31 ›› Issue (2): 205-218.DOI: 10.17521/cjpe.2007.0025
吕超群1,4, 田汉勤2,*( ), 黄耀3
), 黄耀3
收稿日期:2006-11-15
接受日期:2006-12-17
出版日期:2007-11-15
发布日期:2007-03-30
通讯作者:
田汉勤
作者简介:* E mail: tianhan@auburn.edu基金资助:
LÁ Chao-Qun1,4, TIAN Han-Qin2,*(), HUANG Yao3
Received:2006-11-15
Accepted:2006-12-17
Online:2007-11-15
Published:2007-03-30
Contact:
TIAN Han-Qin
摘要:
人类活动在全球范围内极大地改变着氮素从大气向陆地生态系统输入的方式和速率,人为固定的氮素正在不断积累,并对生态系统的结构和功能产生显著影响。该文从以下几个方面综述了大气氮沉降增加对陆地生态系统的影响:1)氮输入增加可能影响植物生产力和生态系统碳蓄积能力,生态系统响应的方向和程度取决于系统的初始氮状况(氮限制或氮饱和)以及当地的植被和土壤特征;2)持续氮输入有可能改变土壤氮循环过程,降低土壤固持氮的能力,甚至导致土壤酸化、盐基离子损耗,进而影响到土壤有机碳的分解;3)高的氮沉降速率和持续氮输入都可能加速含氮痕量气体的释放,但其影响程度受生态系统初始状态的影响(例如磷限制和氮限制);4)氮沉降增加会影响生态系统的物种丰富度、植物群落结构和动态,促进森林扩张,改变菌根真菌的物种多样性;5)持续氮输入带来的植物群落结构和植物生理特征的变化可能影响昆虫取食特性,进而通过食物链改变生态系统的营养结构;6)氮沉降增加对生态系统的影响并不是孤立存在的,它与CO2浓度升高和O3浓度变化有协同作用,但难以从其协同效应中区分出各自的影响。最后,该文总结了我国的氮沉降研究现状,并对今后的研究前景提出了展望。
吕超群, 田汉勤, 黄耀. 陆地生态系统氮沉降增加的生态效应. 植物生态学报, 2007, 31(2): 205-218. DOI: 10.17521/cjpe.2007.0025
LÁ Chao-Qun, TIAN Han-Qin, HUANG Yao. ECOLOGICAL EFFECTS OF INCREASED NITROGEN DEPOSITION IN TERRESTRIAL ECOSYSTEMS. Chinese Journal of Plant Ecology, 2007, 31(2): 205-218. DOI: 10.17521/cjpe.2007.0025
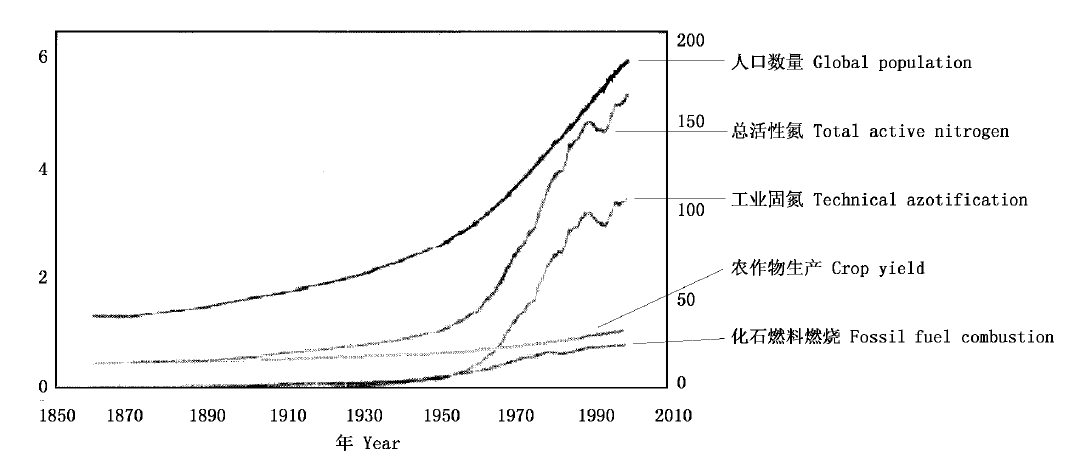
图1 1860~2000年全球活性氮变化趋势,左轴为人口数量(109),右轴为活性氮数量(Tg N·a-1)(Galloway et al., 2003)
Fig.1 Global population trends from 1860 to 2000 (billions, left axis) and reactive nitrogen creation(Tg N per year, right axis)(Galloway et al., 2003)
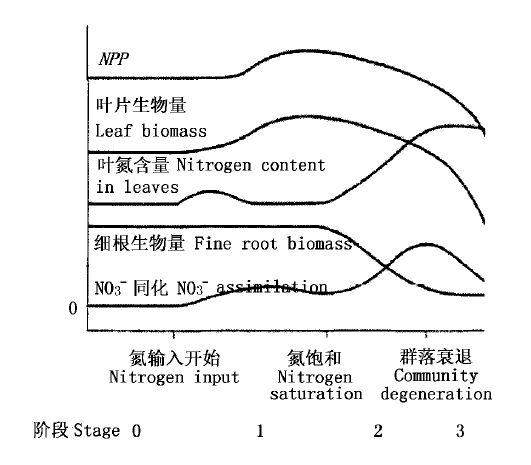
图2 氮限制生态系统对于氮增加的响应(Aber et al., 1989)
Fig.2 The integrated responses of N-limited ecosystems to chronic nitrogen addition (Aber et al., 1989)
| [1] | Aber JD, McDowell W, Nadelhoffer K, Magill A, Berntson G, Kamakea M, McNulty S, Currie W, Rustad L, Fernandez I (1998). Nitrogen saturation in temperate forest ecosystems: hypotheses revisited. BioScience, 48,921-934. |
| [2] | Aber JD, Nadelhoffer KJ, Steudler P, Melillo JM (1989). Nitrogen saturation in northern forest ecosystems. BioScience, 39,378-386. |
| [3] | Asner GP, Townsend AR, Riley WJ, Matson PA, Neff JC, Cleveland CC (2001). Modeling tropical and semi-arid ecosystem responses to increased N deposition. Biogeochemistry, 54,1-39. |
| [4] | Asner GP, Seastedt TR, Townsend AR (1997). The decoupling of terrestrial carbon and nitrogen cycles. BioScience, 47,226-234. |
| [5] | Berg B, Matzner E (1997). Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environmental Reviews, 5,1-25. |
| [6] | Bert D, Leavitt SW, Dupouey JL (1997). Variation of wood δ 13C and water-use efficiency of Abies alba during the last century. Ecology, 78,1588-1596. |
| [7] | Bobbink R, Hornung M, Roelofs JGM (1998). The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. Journal of Ecology, 86,717-738. |
| [8] | Bowden RD, Davidson E, Savage K, Arabia C, Steudler P (2004). Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. Forest Ecology and Management, 196,43-56. |
| [9] | Bowman WD, Theodose TA, Schardt JC, Conant RT (1993). Constraints of nutrient availability on primary production in two alpine tundra communities. Ecology, 74,2085-2097. |
| [10] | Boxman AW, van der Ven PJM, Roelofs JGM (1998). Ecosystem recovery after a decrease in nitrogen input to a Scots pine stand at Ysselsteyn, the Netherlands. Forest Ecology and Management, 101,155-163. |
| [11] | Carreira JA, Harrison AF, Sheppard LJ, Woods C (1997). Reduced soil P availability in a Sitka spruce ( Picea sitchensis (Bong.) Carr.) plantation induced by applied acid-mist: significance in forest decline. Forest Ecology and Management, 92,153-166. |
| [12] |
Casperson JP, Pacala SW, Jenkins JC, Hurtt GC, Moorcroft PR, Birdsey RA (2000). Contributions of land use history to carbon accumulation in U.S. forests. Science, 290,1148-1151.
URL PMID |
| [13] |
Coley PD, Bryant JP, Chapin FS (1985). Resource availability and plant antiherbivore defense. Science, 230,895-899.
DOI URL PMID |
| [14] | Currie WS, Nadelhoffer K (1999). Dynamic redistribution of isotopically labeled cohorts of nitrogen inputs in two temperate forests. Ecosystems, 2,4-18. |
| [15] | Currie WS (1999). The responsive C and N biogeochemistry of the temperate forest floor. Trends in Ecology and Evolution, 14,316-320. |
| [16] | Davidson EA, Kingerlee W (1997). A global inventory of nitric oxide emissions from soils. Nutrient Cycling in Agroecosystem, 48,37-50. |
| [17] | Ding GA (丁国安), Xu XB (徐晓斌), Wang SF (王淑凤), Yu XL (于晓岚), Cheng HB (程红兵) (2004). Database from the acid rain network of China meteorological administration and its preliminary analysis. Journal of Applied Meteorological Science (应用气象学报), 15 (Suppl.),85-94. (in Chinese with English abstract) |
| [18] | Emmett BA, Boxman D, Bredemeier M, Gundersen P, KjØnaas OJ, Moldan F, Schleppi P, Tietema A, Wright RF (1998). Predicting the effects of atmospheric nitrogen deposition in conifer stands: evidence from NITREX ecosystem-scale experiments. Ecosystems, 1,352-360. |
| [19] | Fan S, Gloor M, Mahlman J, Pacala S, Sarmiento J, Takahashi T, Tan P (1998). A large terrestrial carbon sink in North America implied by atmospheric and oceanic carbon dioxide data and models. Science, 282,442-446. |
| [20] | Fang YT (方运霆), Mo JM (莫江明), Zhou GY (周国逸), Gundersen P, Li DJ (李德军), Jiang YQ (江远清) (2004). The short-term responses of soil available nitrogen of Dinghushan forests to simulated N deposition in subtropical China. Acta Ecologica Sinica (生态学报), 24,2353-2359. (in Chinese with English abstract) |
| [21] | Fang YT (方运霆), Mo JM (莫江明), Jiang YQ (江远清), Li DJ (李德军), Gundersen P (2005a). Acidity and inorganic nitrogen concentration in soil solution in short-term response to N addition in subtropical forests. Journal of Tropical and Subtropical Botany (热带亚热带植物学报), 13,123-129. (in Chinese with English abstract) |
| [22] | Fang YT (方运霆), Mo JM (莫江明), Zhou GY(周国逸) (2005b). Response of soil N3--N dynamics to N additions in Dinghushan forest, assessed by ion-exchange resin bag method. Ecology and Environment (生态环境), 14, 483-487. (in Chinese with English abstract) |
| [23] | Fang YT (方运霆), Mo JM (莫江明), Zhou GY (周国逸), Xue JH (薛王景花) (2005c). Response of diameter at breast height increment to N additions in forests of Dinghushan biosphere reserve. Journal of Tropical and Subtropical Botany (热带亚热带植物学报), 13,198-204. (in Chinese with English abstract) |
| [24] | FAO (1993). Food and Agriculture Production Yearbook 1992 . Statistical Series 112.FAO, Rome, Italy. |
| [25] | Fenn ME, Baron JS, Allen EB, Rueth HM, Nydick KR, Geiser L, Bowman WD, Sickman JO, Meixner T, Johnson DW, Neitlich P (2003). Ecological effects of nitrogen deposition in the western United States. BioScience, 53,404-419. |
| [26] | Fenn ME, Poth MA, Johnson DW (1996). Evidence for nitrogen saturation in the San Bernardino Mountains in southern California. Forest Ecology and Management, 82,211-230. |
| [27] | Field CB, Chapin FS, Matson PA III, Mooney HA (1992). Responses of terrestrial ecosystems to the changing atmosphere: a resource-based approach. Annual Review of Ecology and Systematics, 23,201-235. |
| [28] | Firestone MK, Davidson EA (1989). Microbiological basis of NO and N2O production and consumption in soil. In: Andreae MO, Schimel DS eds. Exchange of Trace Gases Between Terrestrial Ecosystems and the Atmosphere. John Wiley & Sons, New York, 7-24. |
| [29] | Galloway JN, Aber JD, Erisman JW, Seitzinger SP, Howarth RW, Cowling EB, Cosby BJ (2003). The nitrogen cascade. BioScience, 53,341-356. |
| [30] | Galloway JN, Levy H II, Kasibhatla PS (1994). Year 2020: consequences of population growth and development on deposition of oxidized nitrogen. Ambio, 23,120-123. |
| [31] | Galloway JN, Schlesinger WH, Levy H II, Michaels A, Schnoor JL (1995). Nitrogen fixation: anthropogenic enhancement-environmental response. Global Biogeochemical Cycles, 9,235-252. |
| [32] | Gifford RM, Lutze JL, Barrett D (1996). Global atmospheric change effects on terrestrial carbon sequestration: exploration with a global C- and N-cycle model (CQUESTN). Plant and Soil, 187,369-387. |
| [33] | Grulker NE, Anderson CP, Fenn ME, Miller PR (1998). Ozone and nitrogen deposition reduces root biomass of ponderosa pine in the San Bernardino Mountains, California. Environment Pollution, 103,63-73. |
| [34] | Gundersen P, Emmett BA, KjØnaas OJ, Koopmans C, Tietema A (1998). Impacts of nitrogen deposition on N cycling: a synthesis of NITREX data. Forest Ecology and Management, 101,37-55. |
| [35] |
Haddad NM, Haarstad J, Tilman D (2000). The effects of long-term nitrogen loading on grassland insect communities. Oecologia, 124,73-84.
URL PMID |
| [36] | Hall SJ, Matson PA (1999). Nitrogen oxide emissions after nitrogen additions in tropical forests. Nature, 400,152-155. |
| [37] | Holland EA, Braswell BH, Lamarque JF, Towsend A, Sulzman J, Müller JF, Dentener F, Brasseur G, Levy H II, Penner JE, Roelofs GJ (1997). Variations in the predicted spatial distribution of atmospheric nitrogen deposition and their impact on carbon uptake by terrestrial ecosystems. Journal of Geophysical Research, 102,849-866. |
| [38] | Holland EA, Dentener FJ, Braswell BH, Sulzman JM (1999). Contemporary and pre-industrial global reactive nitrogen budgets. Biogeochemistry, 46,7-43. |
| [39] | Huang Y (黄耀), Yang ZF (杨兆芳), Yu YQ (于永强), Jiang J (姜军), Sun WJ (孙文娟) (2005). Simulating net primary production of rice and wheat crops: model establishment. Environmental Science (环境科学), 26(2),11-15. (in Chinese with English abstract) |
| [40] | Huang ZL (黄忠良), Kong GH (孔国辉), Yu QF (余清发), Zhang ZH (张志红) (2000). Hydrological function and nutrient dynamics in lower subtropical monsoon evergreen broad-leaved forest. Acta Phytoecologica Sinica (植物生态学报), 24,157-161. (in Chinese with English abstract) |
| [41] | Hudson RJM, Gherini SA, Goldstein RA (1994). Modeling the global carbon cycle: nitrogen fertilization of the terrestrial biosphere and the “missing” CO2 sink. Global Biogeochemical Cycles, 8,307-333. |
| [42] | Huenneke LF, Hamburg ST, Koide R, Mooney HA, Vitousek PM (1990). Effects of soil resources on plant invasion and community structure in Californian serpentine grassland. Ecology, 71,478-491. |
| [43] | Jenkinson DS, Goudling K, Powlson DS, Sievering H, Nadelhoffer KJ (1999). Nitrogen deposition and carbon sequestration. Nature, 400,629-630. |
| [44] | Johnson DW (1992). Nitrogen retention in forest soils. Journal of Environmental Quality, 21,1-12. |
| [45] | Køchy M, Wilson SD (2001). Nitrogen deposition and forest expansion in the northern Great Plains. Journal of Ecology, 89,807-817. |
| [46] | Larssen T, Seip HM, Semb A, Mulder J, Muniz IP, Vogt RD Lydersen E, Angell V, Dagang T, Eilertsen O (1999). Acid deposition and its effects in China: an overview. Environmental Science and Policy, 2,9-24. |
| [47] | Li DJ (李德军), Mo JM (莫江明), Fang YT (方运霆), Cai XA (蔡锡安), Xue JH (薛王景花), Xu GL (徐国良) (2004). Effects of simulated nitrogen deposition on growth and photosynthesis of Schima superba, Castanopsis chinensis and Cryptocarya concinna seedlings. Acta Ecologica Sinica (生态学报), 24,876-882. (in Chinese with English abstract) |
| [48] | Li DJ (李德军), Mo JM (莫江明), Fang YT (方运霆), Li ZA (李志安) (2005a). Effects of simulated nitrogen deposition on biomass production and allocation in Schima superba and Cryptocarya concinna seedlings in subtropical China. Acta Phytoecologica Sinica (植物生态学报), 29,543-549. (in Chinese with English abstract) |
| [49] | Li DJ (李德军), Mo JM (莫江明), Peng SL (彭少麟), Fang YT (方运霆) (2005b). Effects of simulated nitrogen deposition on elemental concentrations of Schima superba and Cryptocarya concinna seedlings in subtropical China. Acta Ecologica Sinica (生态学报), 25,2165-2172. (in Chinese with English abstract) |
| [50] | Lilleskov EA, Fahey TJ, Horton TR, Lovett GM (2002). Belowground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology, 83,103-115. |
| [51] | Liu JX, Price DT, Chen JM (2005). Nitrogen controls on ecosystem carbon sequestration: a model implementation and application to Sakatchewan, Canada. Ecological Modelling, 186,178-195. |
| [52] | Ma XH (马雪华) (1989). Effects of rainfall on the nutrient cycling in man-made forests of Cunninghamia lanceolata and Pinus massoniana. Acta Ecologica Sinica (生态学报), 9,15-20. (in Chinese with English abstract) |
| [53] | Magill AH, Aber JD, Berntson GM, McDowell WH, Nadelhoffer KJ, Melillo JM, Steudler PA (2000). Long-term nitrogen additions and nitrogen saturation in two temperate forests. Ecosystems, 3,238-253. |
| [54] | Magill AH, Aber JD, Currie WS, Nadelhoffer KJ, Martin ME, McDowell WH, Melillo JM, Steudler PA (2004). Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. Forest Ecology and Management, 196,7-28. |
| [55] |
Matson PA, Lohse KA, Hall SJ (2002). The globalization of nitrogen deposition: consequences for terrestrial ecosystems. Ambio, 31,113-118.
DOI URL PMID |
| [56] | Matson PA, McDowell WH, Townsend AR, Vitousek PM (1999). The globalization of N deposition: ecosystem consequences in tropical environments. Biogeochemistry, 46,67-83. |
| [57] | McNulty SG, Aber JD, McLellan TM, Katt SM (1990). Nitrogen cycling in high elevation forests of the Northeastern US in relation to nitrogen deposition. Ambio, 19,38-40. |
| [58] | Melillo JM, Gosz JR (1983). Interaction of biogeochemical cycles in forest ecosystems. In: Bolin B, Cook RB eds. The Major Biogeochemical Cycles and Their Interactions. John Wiley & Sons, Chichester, UK, 177-222. |
| [59] | Mo JM (莫江明), Xue JH (薛王景花), Fang YT (方运霆) (2004). Litter decomposition and its responses to simulated N deposition for the major plants of Dinghushan forests in subtropical China. Acta Ecologica Sinica (生态学报), 24,1413-1420. (in Chinese with English abstract) |
| [60] | Mo JM (莫江明), Fang YT (方运霆), Xu GL (徐国良), Li DJ (李德军), Xue JH (薛王景花) (2005). The short-term responses of soil CO2 emission and CH4 uptake to simulated N deposition in nursery and forests of Dinghushan in subtropical China. Acta Ecologica Sinica (生态学报), 25,682-670. (in Chinese with English abstract) |
| [61] | Nadelhoffer KJ, Emmet BA, Gundersen P, KjØnaas OJ, Koopmans CJ, Schleppi P, Tietemal A, Wright RF (1999). Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests. Nature, 398,145-148. |
| [62] | Neff JC, Bowman WD, Holland EA, Fisk MC, Schmidt SK (1994). Fluxes of nitrous oxide and methane from nitrogen-amended soils in Colorado alpine ecosystem. Biogeochemistry, 27,23-33. |
| [63] |
Neff JC, Townsend AR, Gleixner G, Lehman SJ, Turnbull J, Bowman WD (2002). Variable effects of nitrogen additions on the stability and turnover of organic carbon. Nature, 419,915-917.
DOI URL PMID |
| [64] | Norby RJ (1998). Nitrogen deposition: a component of global change analyses. New Phytologist, 139,189-200. |
| [65] |
Nordin A, Strengbom J, Witzell J, Näsholm T, Ericson L (2005). Nitrogen deposition and the biodiversity of boreal forests: implications for the nitrogen critical load. Ambio, 34,20-24.
URL PMID |
| [66] | Ollinger SV, Aber J, Reich PB, Freuder RJ. (2002). Interactive effects of nitrogen deposition, tropospheric ozone, elevated CO2 and land use history on the carbon dynamics of northern hardwood forests. Global Change Biology, 8,545-562. |
| [67] | Read DJ (1991). Mycorrhizas in ecosystems—nature's response to the “law of the minimum”. In: Hawksworth DL ed. Frontiers in Mycology. CAB International, Regensburg, Germany,101-130. |
| [68] | Schimel DS, Alevs D, Enting I, Heimann M (1996). Radiative forcing of climate change. In: Houghton JT, Meira FLG, Callender BA, Harris N, Kattenberg A, Maskell K eds. Climate Change 1995: the Science of Climate Change. IPCC WG 1 Report. Cambridge University Press, Cambridge, UK,65-131. |
| [69] | Schindler DW, Bayley SE (1993). The biosphere as an increasing sink for atmospheric carbon: estimates from increased nitrogen deposition. Global Biogeochemical Cycles, 7,717-733. |
| [70] | Schleppi P, Bucher-Wallin I, Siegwolf R, Saurer M, Muller N, Bucher JB (1999). Simulation of increased nitrogen deposition to a montane forest ecosystem: partitioning of the added 15N. Water, Air, & Soil Pollution, 116,129-134. |
| [71] | Siemann E (1998). Experimental tests of the effects of plant productivity and plant diversity on grassland arthropod diversity. Ecology, 79,2057-2070. |
| [72] | Streets DG, Bond TC, Carmichael GR, Fernandes SD, Fu Q, He D, Klimont Z, Nelson SM, Tsai NY, Wang MQ, Woo JH, Yarber KF (2003). An inventory of gaseous and primary aerosol emissions in Asia in the year 2000. Journal of Geographical Research, 108(D21),8809, doi:10.1029/2002JD003093. |
| [73] | Sun QR (孙庆瑞), Wang MR (王美蓉) (1997). Ammonia emission and concentration in the atmosphere over China. Chinese Journal of Atmospheric Sciences (大气科学), 21,590-598. (in Chinese with English abstract) |
| [74] | Takemoto BK, Bytnerowicz A, Fenn ME (2001). Current and future effects of ozone and atmospheric nitrogen on California's mixed conifer forests. Forest Ecology and Management, 144,159-173. |
| [75] | Throop HL, Holland EA, Parton WJ, Ojima DS, Keough CA (2004). Effects of nitrogen deposition and insect herbivory on patterns of ecosystem-level carbon and nitrogen dynamics: results from the CUNTURY model. Global Change Biology, 10,1092-1105. |
| [76] | Tian DL (田大伦), Pan WC (潘维俦), Chen XY (陈小勇), Zhu XN (朱小年) (1994). Studies of biogeochemical cycling in Chinese fir plantation ecosystems. In: Zhou XF (周晓峰) ed. Long-Term Research on China's Forest Ecosystems (森林生态系统定位研究). Northeast Forestry University Press, Harbin, 136-145. (in Chinese) |
| [77] | Tian HQ, Hall CAS, Qi Y (1998). Modeling primary productivity of the terrestrial biosphere in changing environments: toward a dynamic biosphere model. Critical Review in Plant Sciences, 17,541-557. |
| [78] | Tian HQ, Melillo JM, Kicklighter DW, Pan S, Liu J, McGuire AD, Moore B III (2003). Regional carbon dynamics in monsoon Asia and its implications to the global carbon cycle. Global and Planetary Change, 37,201-217. |
| [79] | Tian HQ, Wang S, Liu J, Pan S, Chen H, Zhang C, Shi X (2006). Patterns of soil nitrogen storage in China. Global Biogeochemical Cycles, 20, GB1001, doi:10.1029/2005GB002464. |
| [80] | Tietema A, Emmett BA, Gundersen P, KjØnaas OJ, Koopmans CJ (1998). The fate of 15N-labelled nitrogen deposition in coniferous forest ecosystem. Forest Ecology and Management, 101,19-27. |
| [81] | Townsend AR, Braswell BH, Holland EA, Penner JE (1996). Spatial and temporal patterns in terrestrial carbon storage due to deposition of anthropogenic nitrogen. Ecological Applications, 6,806-814. |
| [82] | Treseder KK (2004). A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytologist, 164,347-355. |
| [83] | Uehara G, Gillman G (1981). The Mineralogy, Chemistry and Physics of Tropical Soils with Variable Charge Clays. Westview Press, Boulder, Colorado, 160. |
| [84] | Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler DW, Schlesinger WH, Tilman GD (1997). Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications, 7,737-750. |
| [85] | Wang WX (王文兴), Wang W (王玮), Zhang WH (张婉华), Hong SX (洪少贤) (1996). Geographical distribution of SO2 and NOx emission intensities and trends in China. China Environmental Science (中国环境科学), 16,161-167. (in Chinese with English abstract) |
| [86] |
Wedin DA, Tilman D (1996). Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science, 274,1720-1723.
DOI URL PMID |
| [87] | Wilson SD (1998). Competition between grasses and woody plants. In: Cheplick GP ed. Population Ecology of Grasses. Cambridge University Press, Cambridge, UK, 231-254. |
| [88] | Wilson SD, Tilman D (1991). Components of plant competition along an experimental gradient of nitrogen availability. Ecology, 72,1050-1065. |
| [89] | Xu GL (徐国良), Mo JM (莫江明), Sandra B, Li DJ (李德军) (2004). Effects of simulated N deposition on soil fauna. Acta Ecologica Sinica (生态学报), 24,2245-2251. (in Chinese with English abstract) |
| [90] | Xu GL (徐国良), Mo JM (莫江明), Zhou GY (周国逸), Xue JH (薛王景花) (2005). Litter decomposition under N deposition in Dinghushan forests and its relationship with soil fauna. Ecology and Environment (生态环境), 14,901-907. (in Chinese with English abstract) |
| [1] | 俞庆水 倪晓凤 吉成均 朱江玲 唐志尧 方精云. 10年氮磷添加对海南尖峰岭两种热带雨林优势植物叶片非结构性碳水化合物的影响[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 陈保冬, 付伟, 伍松林, 朱永官. 菌根真菌在陆地生态系统碳循环中的作用[J]. 植物生态学报, 2024, 48(1): 1-20. |
| [3] | 张英, 张常洪, 汪其同, 朱晓敏, 尹华军. 氮沉降下西南山地针叶林根际和非根际土壤固碳贡献差异[J]. 植物生态学报, 2023, 47(9): 1234-1244. |
| [4] | 仲琦, 李曾燕, 马炜, 况雨潇, 邱岭军, 黎蕴洁, 涂利华. 氮添加和凋落物处理对华西雨屏区常绿阔叶林凋落叶分解的影响[J]. 植物生态学报, 2023, 47(5): 629-643. |
| [5] | 冯继广, 张秋芳, 袁霞, 朱彪. 氮磷添加对土壤有机碳的影响: 进展与展望[J]. 植物生态学报, 2022, 46(8): 855-870. |
| [6] | 张英, 张常洪, 汪其同, 朱晓敏, 尹华军. 氮沉降下西南山地针叶林根际和非根际土壤微生物养分限制特征差异[J]. 植物生态学报, 2022, 46(4): 473-483. |
| [7] | 田磊, 朱毅, 李欣, 韩国栋, 任海燕. 不同降水条件下内蒙古荒漠草原主要植物物候对长期增温和氮添加的响应[J]. 植物生态学报, 2022, 46(3): 290-299. |
| [8] | 谢欢, 张秋芳, 曾泉鑫, 周嘉聪, 马亚培, 吴玥, 刘苑苑, 林惠瑛, 尹云锋, 陈岳民. 氮添加对杉木苗期磷转化和分解类真菌的影响[J]. 植物生态学报, 2022, 46(2): 220-231. |
| [9] | 吴霖升, 张永光, 章钊颖, 张小康, 吴云飞. 日光诱导叶绿素荧光遥感及其在陆地生态系统监测中的应用[J]. 植物生态学报, 2022, 46(10): 1167-1199. |
| [10] | 朱湾湾, 王攀, 许艺馨, 李春环, 余海龙, 黄菊莹. 降水量变化与氮添加下荒漠草原土壤酶活性及其影响因素[J]. 植物生态学报, 2021, 45(3): 309-320. |
| [11] | 魏春雪, 杨璐, 汪金松, 杨家明, 史嘉炜, 田大栓, 周青平, 牛书丽. 实验增温对陆地生态系统根系生物量的影响[J]. 植物生态学报, 2021, 45(11): 1203-1212. |
| [12] | 张宏锦, 王娓. 生态系统多功能性对全球变化的响应: 进展、问题与展望[J]. 植物生态学报, 2021, 45(10): 1112-1126. |
| [13] | 庞芳, 夏维康, 何敏, 祁珊珊, 戴志聪, 杜道林. 固氮菌缓解氮限制环境中丛枝菌根真菌对加拿大一枝黄花的营养竞争[J]. 植物生态学报, 2020, 44(7): 782-790. |
| [14] | 冯继广, 朱彪. 氮磷添加对树木生长和森林生产力影响的研究进展[J]. 植物生态学报, 2020, 44(6): 583-597. |
| [15] | 牛书丽, 陈卫楠. 全球变化与生态系统研究现状与展望[J]. 植物生态学报, 2020, 44(5): 449-460. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19