
植物生态学报 ›› 2005, Vol. 29 ›› Issue (3): 487-496.DOI: 10.17521/cjpe.2005.0065
王永峰1,2, 李庆军1,*( )
)
收稿日期:2004-06-02
接受日期:2004-12-14
出版日期:2005-05-30
发布日期:2005-05-30
通讯作者:
李庆军
作者简介:*E-mail: qjlixtbg@bn.yn.cninfo.net基金资助:
WANG Yong-Feng1,2, LI Qing-Jun1,*()
Received:2004-06-02
Accepted:2004-12-14
Online:2005-05-30
Published:2005-05-30
Contact:
LI Qing-Jun
摘要:
综述了国内外生物源挥发性有机化合物 (Biologicalvolatileorganiccompounds, BVOCs) 研究现状及未来的研究方向, 侧重介绍了陆地生态系统中植物排放BVOCs的种类、生物学功能及其对大气化学过程的影响。BVOCs按其化学结构以及在大气中的滞留时间可以分为 4类 :异戊二烯、单萜、其它活性BVOCs和其它次活性BVOCs。不同的植物类群排放不同的BVOCs种类并具有不同的排放特性, 环境条件对植物不同BVOCs的排放影响也不同。BVOCs作为有机物质被排放到体外, 从植物能量代谢的角度来讲要消耗一部分植物光合作用产物从而降低植物的生产力, 因此推测植物排放BVOCs具有一定的生理学或者生态学的功能。其中比较成熟的假说是抗热胁迫假说, 其次是抗氧化假说, 也有一些其它假说例如促氮同化假说等。但这些假说都还缺乏直接的有力证据, 有待更多的研究来支持。BVOCs被排放到大气中对大气化学过程的影响更是科学家关注的问题, BVOCs对大气的影响一方面是在大气对流层中促进臭氧 (O3 ) 的形成, 造成环境污染, 另一方面BVOCs通过对大气中的OH自由基和臭氧等氧化物浓度的调整而影响到大气中甲烷等温室气体的平衡, 对大气温室效应具有间接的贡献。我国在BVOCs的研究上也做了大量的工作, 包括分析鉴定了一些植物排放的BVOCs, 探讨了环境因子对植物BVOCs排放速率的影响, 从不同尺度估测了BVOCs的排放量等等。今后对BVOCs的研究将会集中在以下几个方面 :1) 进一步研究不同植物类群释放的BVOCs种类及其它们在大气中的理化性质 ;2 ) 继续探讨植物排放BVOCs的合成与代谢途径及其生物学功能 ;3) 研究BVOCs对大气化学过程的作用, 以及区域植被变化对BVOCs排放格局进而对区域乃至全球环境变化的影响 ;4 ) 加强对一些研究比较薄弱的生态系统例如在热带地区所进行的BVOCs研究工作 ;5 ) 进一步建立和完善BVOCs排放的理论模型, 以模拟不同陆地生态系统BVOCs排放的时空动态。
王永峰, 李庆军. 陆地生态系统植物挥发性有机化合物的排放及其生态学功能研究进展. 植物生态学报, 2005, 29(3): 487-496. DOI: 10.17521/cjpe.2005.0065
WANG Yong-Feng, LI Qing-Jun. BVOCs EMITTED FROM PLANTS OF TERRESTRIAL ECOSYSTEMSAND THEIR ECOLOGICAL FUNCTIONS. Chinese Journal of Plant Ecology, 2005, 29(3): 487-496. DOI: 10.17521/cjpe.2005.0065
| 种类 Species | 在大气中的寿命 Lifetime in air (h) | 化学式 Formula | 全球年排放量 Global emission (Tg C·a-1) | 主要代表物 Example |
|---|---|---|---|---|
| 异戊二烯 Isoprene | 1~2 | C5H8 | 503 | 异戊二烯 Isoprene |
| 单萜 Monoterpene | 0.5~3 | C10H18 | 127 | α-蒎烯, β-蒎烯 α- pinene, β-pinene |
| 其它活性BVOCs Other reactive BVOCs | < 24 | CxHyOz | 260 | 甲基丁烯醇, 己烯醛 Methylbutenol, Hexenal |
| 其它次活性BVOCs Other less reactive BVOCs | > 24 | CxHyOz | 260 | 甲醇, 乙醇, 丙酮 Methanol, Ethanol, Acetone |
表1 生物源挥发性有机化合物 (BVOCs) 的种类及其特性
Table 1 Categories and traits of biogenic volatile organic compounds (BVOCs)
| 种类 Species | 在大气中的寿命 Lifetime in air (h) | 化学式 Formula | 全球年排放量 Global emission (Tg C·a-1) | 主要代表物 Example |
|---|---|---|---|---|
| 异戊二烯 Isoprene | 1~2 | C5H8 | 503 | 异戊二烯 Isoprene |
| 单萜 Monoterpene | 0.5~3 | C10H18 | 127 | α-蒎烯, β-蒎烯 α- pinene, β-pinene |
| 其它活性BVOCs Other reactive BVOCs | < 24 | CxHyOz | 260 | 甲基丁烯醇, 己烯醛 Methylbutenol, Hexenal |
| 其它次活性BVOCs Other less reactive BVOCs | > 24 | CxHyOz | 260 | 甲醇, 乙醇, 丙酮 Methanol, Ethanol, Acetone |
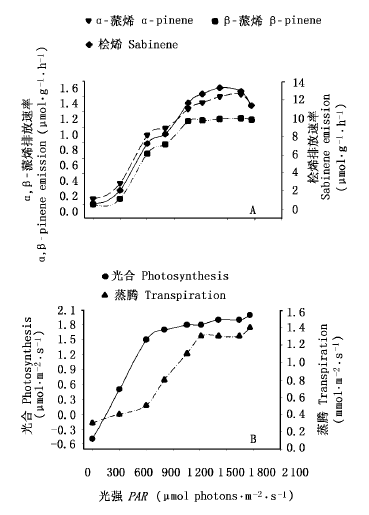
图1 光强对橡胶排放α-蒎烯, β-蒎烯和桧烯的影响
Fig.1 Effect of increasing photosynthesis active radiation (PAR) on α-pinene, sabinene and β-pinene emissions of rubber tree (Hevea brasiliensis)
| [1] |
Affek HP, Yakir D (2002). Protection by isoprene against singlet oxygen in leaves. Plant Physiology, 129,269-277.
DOI URL |
| [2] | Asada K (1994). Production and action of active oxygen species in photosynthetic tissues. In: Foyer CH, Mullineaux PM eds. Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants. CRC, London,77-104. |
| [3] | Bai JH (白建辉), Baker B (2004). Model simulation of isoprene emission flux in a tropical forest plantation of rubber trees. Acta Scientiae Circumstantiae (环境科学学报), 24,197-203. (in Chinese with English abstract) |
| [4] | Bai JH (白建辉), Wang GC (王庚辰), Ren LX (任丽新), Baker B, Zimmerman P, Liang BS (梁宝生) (2003). The emission flux of volatile organic compounds in the Inner Mongolia grassland. Environmental Science (环境科学), 24,17-22. (in Chinese with English abstract) |
| [5] | Bai JH (白建辉), Wang MX (王明星), Graham JR, Prinn G (2001). A study of the nonmethane hydrocarbons at subtropical forest Part Ⅱ. Diurnal variation. Climatic and Environmental Research (气候与环境研究), 6,456-466. (in Chinese with English abstract) |
| [6] | Bai JH (白建辉), Wang MX (王明星), Graham J, Prinn RG, Huang ZL (黄忠良) (1998). Primary study on the concentrations of nonmethane hydrocarbon emitted from the forest. Scientia Atmospherica Sinica (大气科学), 22,247-251. (in Chinese with English abstract) |
| [7] | Bai YH (白郁华), Li JL (李金龙), Zhao MP (赵美萍), Xia Y (夏羽), Tang XY (唐孝炎) (1995). Relative factor of hydrocarbons emission from popular tree. Environmental Chemistry (环境化学), 14,118-123. (in Chinese with English abstract) |
| [8] |
Benjamin MT (1996). Low-emitting urban forests: a taxonomic methodology for assessing isoprene and monoterpene emission rates. Atmospheric Environment, 30,1437-1452.
DOI URL |
| [9] | Brasseur G, Chatfield R (1991). The fate of biogenic trace gases in the atmosphere. In: Sharkey T, Holland E, Mooney H eds. Trace Gas Emissions by Plants. Academic Press, San Diego,1-27. |
| [10] |
Carter WPL, Atkinson R (1996). Development and evaluation of a detailed mechanism for the atmospheric reactions of isoprene and NOx. International Journal of Chemical Kinetics, 28,497-530.
DOI URL |
| [11] |
Delwiche CF, Sharkey TD (1993). Rapid appearance of 13C in biogenic isoprene when 13CO2 is fed to intact leaves. Plant, Cell and Environment, 16,587-591.
DOI URL |
| [12] |
De Moraes CM, Mescher MC, Tumlinson JH (2001). Caterpillar-induced nocturnal plant volatiles repel conspecific females. Nature, 410,577-580.
PMID |
| [13] | Fehsenfeld F, Calvert J, Fall R, Goldan P, Guenther AB, Lamb B, Trainer M, Westberg H, Zimmerman P, Hewitt CN (1992). Emissions of volatile organic compounds from vegetation and the implications for atmospheric chemistry. Global Biogeochemical Cycles, 6,389-430. |
| [14] | Fuentes JD, Hayden BP, Garstang M, Lerdau M, Fitzjarrald D, Baldocchi DD, Monson R, Lamb B, Geron C (2000). VOCs and biosphere-atmosphere feedbacks. Atmospheric Environment, 31,189-191. |
| [15] |
Gershenzon J, McConkey ME, Croteau RB (2000). Regulation of monoterpene accumulation in leaves of peppermint. Plant Physiology, 122,205-213.
DOI URL |
| [16] |
Gray DW, Lerdau MT, Goldstein AH (2003). Influrences of temperature history, water stress, and needle age on methylbutenol emissions. Ecology, 84,765-776.
DOI URL |
| [17] |
Guenther AB, Hewitt CN, Erickson D, Fall R, Geron C, Graedel T, Harley P, Klinger L, Lerdau M, McKay WA, Pierce T, Scholes B, Steinbrecher R, Tallamraju R, Taylor J, Zimmerman P (1995). A global model of natural volatile organic compound emissions. Journal of Geophysical Research, 100,8873-8892.
DOI URL |
| [18] |
Guenther AB, Monson RK, Fall R (1991). Isoprene and monoterpene emission rate variability: observations with eucalyptus and emission rate algorithm development. Journal of Geophysical Research, 96,10799-10808.
DOI URL |
| [19] |
Harley P, Fridd-Stroud V, Greenberg J, Guenther A, Vasconcellos P (1998). Emission of 2-methyl-3-buten-2-ol by pines: a potentially large natural source of reactive carbon to the atmosphere. Journal of Geophysical Research, 103,25479-25486.
DOI URL |
| [20] | Hewitt CN, Kok GL, Fall R (1990). Hydroperoxides in plants exposed to ozone mediate air pollution damage to alkene emitters. Nature, 344,56-58. |
| [21] |
Jacob DJ, Wofsy SC (1988). Photochemistry of biogenic emissions over the Amazon forest. Journal of Geophysical Research, 93,1477-1486.
DOI URL |
| [22] |
Kavouras LG, Mihalopoulos N, Stephanou EG (1998). Formation of atmospheric particles from organic acids produced by forests. Nature, 395,683-686.
DOI URL |
| [23] |
Kesselmeier J, Staudt M (1999). Biogenic volatile organic compounds (VOC) : an overview on emission, physiology and ecology. Journal of Atmospheric Chemistry, 33,23-88.
DOI URL |
| [24] |
Klinger LF, Greenberg JP, Guenther AB, Tyndall G, Zimmerman P, M'Bangui M, Moutsamboté JM, Kenfack D (1998). Patterns in volatile organic compound emissions along a savanna-rainforest gradient in central Africa. Journal of Geophysical Research, 103,1443-1454.
DOI URL |
| [25] | Klinger LF, Li QJ, Guenther AB, Greenberg JP, Baker B, Bai JH (2002). Assessment of volatile organic compound emissions from ecosystems of China. Journal of Geophysical Research, 107,4603-4624. |
| [26] |
Langenheim J (1994). Higher plant terpenoids: a phytocentric overview of their ecological roles. Journal of Chemical Ecology, 20,1223-1280.
DOI URL |
| [27] |
Larom D, Garstang M, Lindeque M, Raspet R, Zunckel M, Hong Y, Brassel K, O'Beirne S, Sokolic F (1997). The influence of surface atmospheric conditions on the range and area reached by animal vocalizations. Journal of Experimental Biology, 200,421-431.
PMID |
| [28] | Li JL (李金龙), Bai YH (白郁华), Hu JX (胡建信), Shao M (邵敏), Zhang BX (张宝祥), Xia Y (夏羽), Tang XY (唐孝炎) (1994). Diurnal variation in the concentration of terpenes and its emission rate measurements from oil pine. China Environmental Science (中国环境科学), 14,165-169. (in Chinese with English abstract) |
| [29] | Li QJ (李庆军), Klinger LF (2001). The correlation between the volatile organic compound emissions and the vegetation succession of ecosystems in different climatic zones of China. Acta Botanica Sinica (植物学报), 43,1065-1071. (in Chinese with English abstract) |
| [30] |
Lichtenthaler HK, Schwender J, Disch A, Rohmer M (1997). Biosynthesis of isoprenoids in higher plant chloroplasts proceeds via a mevalonate-independent pathway. FEBS Letters, 400,271-274.
DOI URL |
| [31] | Loreto F, Ciccioli P, Brancaleoni E, Valentini R, Lillis MD, Csiky O, Seufert G (1998). A hypothesis on the evolution of isoprenoid emission by oaks based on the correlation between emission type and Quercus taxonomy. Oecologia, 11,302-305. |
| [32] |
Loreto F, Ciccioli P, Cecinato A, Brancaleoni E, Frattoni M, Tricoli D (1996). Influence of environmental factors and air composition on the emission of alpha-pinene from Quercus ilex leaves. Plant Physiology, 110,267-275.
PMID |
| [33] |
Loreto F, Mannozzi M, Maris C, Nascetti P, Ferranti F, Pasqualini S (2001). Ozone quenching properties of isoprene and its antioxidant role in leaves. Plant Physiology, 126,993-1000.
DOI URL |
| [34] |
Loreto F, Sharkey TD (1990). A gas-exchange study of photosynthesis and isoprene emission in Quercus rubru L. Planta, 182,523-531.
DOI PMID |
| [35] |
Loreto F, Velikova V (2001). Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiology, 127,1781-1787.
DOI URL |
| [36] |
Martin DM, Gershenzon J, Bohlmann J (2003). Induction of volatile terpene biosynthesis and diurnal emission by Methyl Jasmonate in foliage of Norway spruce. Plant Physiology, 132,1586-1599.
DOI URL |
| [37] |
Martin PH, Guenther AB (1995). Insights into the dynamics of forest succession and non-methane hydrocarbon trace gas emissions. Journal of Biogeography, 22,493-499.
DOI URL |
| [38] |
Monson R, Harley P, Litvak M, Wildermuth M, Guenther A, Zimmerman P, Fall R (1994). Environmental and developmental controls over the seasonal pattern of isoprene emission from aspen leaves. Oecologia, 99,260-270.
DOI PMID |
| [39] |
Niinemets M, Reichstein M, Seufert G, Tenhunen JD (2002). Stomatal constraints may affect emission of oxygenated monoterpenoids from the foliage of Pinus pinea. Plant Physiology, 130,1371-1385.
DOI URL |
| [40] | Owen SM, Harley P, Guenther AB, Hewitt CN (2002). Light dependency of VOC emissions from selected Mediterranean plant species. Atmospheric Environment, 36,3147-3159. |
| [41] |
Paré PW, Tumlinson JH (1997). De novo biosynthesis of volatiles induced by insect herbivory in cotton plants. Plant Physiology, 114,1161-1167.
DOI URL |
| [42] |
Peñuelas J, Llusià J (2001). The complexity of factors driving volatile organic compound emissions by plants. Biologia Plantarum, 44,481-487.
DOI URL |
| [43] |
Peñuelas J, Llusià J (2003). BVOCs: plant defense against climate warming? Trends in Plant Science, 8,105-109.
DOI URL |
| [44] |
Rohmer M (1993). The biosynthesis of triterpenoids of the hopane series in the Eubacteria: a mine of new enzyme reactions. Pure and Applied Chemistry, 65,1293-1298.
DOI URL |
| [45] |
Rohmer M, Knani M, Simonin P, Sahm H (1993). Isoprenoid biosynthesis in bacteria: a novel pathway for the early steps leading to isopentenyl diphosphate. The Biochemical Journal, 295,517-524.
DOI URL |
| [46] |
Rosenstiel TN, Fisher AJ, Fall R, Monson RK (2002). Differential accumulation of dimethylallyl diphosphate in leaves and needles of isoprene- and methylbutenol-emitting and nonemitting species. Plant Physiology, 129,1276-1284.
DOI URL |
| [47] | Sharkey TD, Singsaas EL (1995). Why plants emit isoprene? Nature, 374,769. |
| [48] |
Sharkey TD, Chen X, Yeh S (2001). Isoprene increases thermotolerance of fosmidomycin-fed leaves. Plant Physiology, 125,2001-2006.
DOI URL |
| [49] | Sharkey TD, Yeh S (2001). Isoprene emission from plants. Annual Review of Plant Physiology & Plant Molecular Biology, 52,407-436. |
| [50] |
Shallcross DE, Monks PS (2000). A role for isoprene in biosphere-climate-chemistry feedbacks? Atmospheric Environment, 34,1659-1660.
DOI URL |
| [51] |
Shen B, Jensen RG, Bohnert HJ (1997). Mannitol protects against oxidation by hydroxyl radicals. Plant Physiology, 115,527-532.
PMID |
| [52] |
Singsaas EL, Lerdau M, Winter K, Sharkey TD (1997). Isoprene increases thermotolerance of isoprene-emitting species. Plant Physiology, 115,1413-1420.
PMID |
| [53] |
Staudt M, Seufert G (1995). Light-dependent emissions of monoterpenes by holm oak (Quercus ilex L.). Naturwissenschaften, 82,89-92.
DOI URL |
| [54] |
Terry GM, Stokes NJ, Hewitt CN, Mansfield TA (1995). Exposure to isoprene promotes flowering in plants. Journal of Experimental Botany, 46,1629-1631.
DOI URL |
| [55] | Tingey TD, Turner DP, Weber JA (1991). Factors controlling the emissions of monoterpenes and other volatile organic compounds. Trace Gas Emissions by Plants. Academic Press, San Diego,93-119. |
| [56] |
Trainer M, Williams E, Parrish D, Buhr M, Allwine E, Westberg H, Fehsenfeld F, Liu S (1987). Models and observations of the impact of natural hydrocarbons on rural ozone. Nature, 329,705-707.
DOI URL |
| [57] |
Tuazon EC, Atkinson R (1990). A product study of the gas-phase reaction of isoprene with the OH radical in the presence of NOx. International Journal of Chemical Kinetics, 22,1221-1236.
DOI URL |
| [58] |
Wildermuth MC, Fall R (1998). Biochemical characterization of stromal and thylakoid-bound isoforms of isoprene synthase in willow leaves. Plant Physiology, 116,1111-1123.
PMID |
| [59] | Zhang L (张莉), Bai YY (白艳莹), Wang XK (王效科), Ouyang ZY (欧阳志云), Mu YJ (牟玉静), Miao QL (缪启龙) (2002). Isoprene emission of bamboo and its implication to ozone level in region. Acta Ecologica Sinica (生态学报), 22,1339-1344. (in Chinese with English abstract) |
| [60] | Zhao MP (赵美萍), Shao M (邵敏), Bai YH (白郁华), Li JL (李金龙), Tang XY (唐孝炎) (1996). Study on NMHC emission characteristics of several typical trees in China. Environmental Chemistry (环境化学), 15,69-75. (in Chinese with English abstract) |
| [1] | 邓蓓 王晓锋 廖君. 环境胁迫影响三峡库区消落带草本和木本植物生理生态特征的整合分析[J]. 植物生态学报, 2024, 48(5): 623-637. |
| [2] | 陈以恒 玉素甫江·如素力 阿卜杜热合曼·吾斯曼. 2001-2020年天山新疆段草地植被覆盖度时空变化及驱动因素分析[J]. 植物生态学报, 2024, 48(5): 561-576. |
| [3] | 张计深, 史新杰, 刘宇诺, 吴阳, 彭守璋. 气候变化下中国潜在自然植被生态系统碳储量动态[J]. 植物生态学报, 2024, 48(4): 428-444. |
| [4] | 臧妙涵, 王传宽, 梁逸娴, 刘逸潇, 上官虹玉, 全先奎. 基于纬度移栽的落叶松叶、枝、根生态化学计量特征对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 469-482. |
| [5] | 梁逸娴, 王传宽, 臧妙涵, 上官虹玉, 刘逸潇, 全先奎. 落叶松径向生长和生物量分配对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 459-468. |
| [6] | 吴茹茹, 刘美珍, 谷仙, 常馨月, 郭立月, 蒋高明, 祁如意. 气候变化对巨柏适宜生境分布的潜在影响和预测[J]. 植物生态学报, 2024, 48(4): 445-458. |
| [7] | 秦文宽, 张秋芳, 敖古凯麟, 朱彪. 土壤有机碳动态对增温的响应及机制研究进展[J]. 植物生态学报, 2024, 48(4): 403-415. |
| [8] | 杨宇萌, 来全, 刘心怡. 气候变化和人类活动对内蒙古植被总初级生产力的定量影响[J]. 植物生态学报, 2024, 48(3): 306-316. |
| [9] | 张启, 程雪寒, 王树芝. 北京西山老龄树记载的森林干扰历史[J]. 植物生态学报, 2024, 48(3): 341-348. |
| [10] | 陈保冬, 付伟, 伍松林, 朱永官. 菌根真菌在陆地生态系统碳循环中的作用[J]. 植物生态学报, 2024, 48(1): 1-20. |
| [11] | 白雨鑫, 苑丹阳, 王兴昌, 刘玉龙, 王晓春. 东北地区3种桦木木质部导管特征对气候变化响应的趋同与差异[J]. 植物生态学报, 2023, 47(8): 1144-1158. |
| [12] | 冯珊珊, 黄春晖, 唐梦云, 蒋维昕, 白天道. 细叶云南松针叶形态和显微性状地理变异及其环境解释[J]. 植物生态学报, 2023, 47(8): 1116-1130. |
| [13] | 黄鹏, 林勇文, 张杰, 姚锦爱, 余德亿. 榕属植物特定挥发物决定榕管蓟马的寄主选择行为[J]. 植物生态学报, 2023, 47(7): 954-966. |
| [14] | 余继梅, 吴福忠, 袁吉, 金遐, 魏舒沅, 袁朝祥, 彭艳, 倪祥银, 岳楷. 全球尺度上凋落物初始酚类含量特征及影响因素[J]. 植物生态学报, 2023, 47(5): 608-617. |
| [15] | 赵小祥, 朱彬彬, 田秋香, 林巧玲, 陈龙, 刘峰. 叶片凋落物分解的主场优势研究进展[J]. 植物生态学报, 2023, 47(5): 597-607. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19