
植物生态学报 ›› 2017, Vol. 41 ›› Issue (5): 559-569.DOI: 10.17521/cjpe.2016.0248
颉洪涛, 虞木奎, 成向荣*( )
)
出版日期:2017-05-10
发布日期:2017-06-22
通讯作者:
成向荣
作者简介:* 通信作者Author for correspondence (E-mail:基金资助:
Hong-Tao XIE, Mu-Kui YU, Xiang-Rong CHENG*()
Online:2017-05-10
Published:2017-06-22
Contact:
Xiang-Rong CHENG
About author:KANG Jing-yao(1991-), E-mail: 摘要:
增强对耐阴植物在不同光环境下氮磷生理响应的理解可以为改善人工林林分结构和提高生态系统功能提供借鉴。该文选取5种亚热带地区具有较高生态和经济价值的耐阴植物幼苗为研究对象, 通过控制实验模拟不同光环境, 设置5个梯度探究光照强度变化对耐阴植物氮磷养分含量、分配以及限制状况的影响。结果显示: 随着光强减弱, 5种耐阴植物的总生物量和总氮磷积累均呈降低趋势, 但各器官氮磷含量有增加趋势。其中, 栀子(Gardenia jasminoides)氮含量最高, 红茴香(Illicium henryi)氮含量最低; 乌冈栎(Quercus phillyraeoides)和栀子的磷含量明显高于山杜英(Elaeocarpus sylvestris)、朱砂根(Ardisia crenata)和红茴香。栀子和乌冈栎在极低光强(6%自然光强)下氮和磷含量均为最高, 朱砂根和红茴香则分别在低光强(15%自然光强)和中等光强(33%自然光强和52%自然光强)时氮磷含量最高; 而杜英氮磷含量最高时对应的光照强度不同, 分别为52%自然光强和6%自然光强。杜英、朱砂根和红茴香各器官的氮磷含量均为叶片>根系>枝干, 栀子和乌冈栎为根系>叶片>枝干。随光强减弱, 氮磷利用格局发生变化。在较高光强(100%自然光强)下, 5种植物各器官氮含量变化表现出良好的一致性, 但在较低光强(15%自然光强和6%自然光强)下表现出明显的分异; 光强减弱时, 植物各器官磷含量变化在红茴香、栀子和乌冈栎上表现出一致性, 但在杜英和朱砂根上表现为分异性。光强减弱显著影响耐阴植物器官氮磷比, 但未根本改变植物生长的元素受限状况。光强变化和树种差异均对耐阴植物在异质性光环境中的氮磷利用和分配产生调节, 进而影响总生物量和总氮磷积累, 这源于植物氮磷利用策略的改变。因此, 在构建复层异龄林时应考虑耐阴植物氮磷养分平衡对光强的偏好。
颉洪涛, 虞木奎, 成向荣. 光照强度变化对5种耐阴植物氮磷养分含量、分配以及限制状况的影响. 植物生态学报, 2017, 41(5): 559-569. DOI: 10.17521/cjpe.2016.0248
Hong-Tao XIE, Mu-Kui YU, Xiang-Rong CHENG. Effects of light intensity variation on nitrogen and phosphorus contents, allocation and limitation in five shade-enduring plants. Chinese Journal of Plant Ecology, 2017, 41(5): 559-569. DOI: 10.17521/cjpe.2016.0248
| 物种 Species | 缩写 Abbreviation | 科属 Family and genera | 生活型 Life form | 主要用途 Main applications |
|---|---|---|---|---|
| 山杜英 Elaeocarpus sylvestris | ES | 杜英科杜英属 Elaeocarpaceae Elaeocarpus | 乔木 Tree | 园林、木材、防护、药用等 Garden, timber, protection, medicine, etc. |
| 朱砂根 Ardisia crenata | AC | 紫金牛科紫金牛属 Myrsinaceae Ardisia | 灌木 Shrub | 观赏、固土、药用等 Ornamental, soil fixing, medicine, etc. |
| 红茴香 Illicium henryi | IH | 八角科八角属 Illiciaceae Illicium | 乔木 Tree | 观赏、药用等 Ornamental, medicine, etc. |
| 栀子 Gardenia jasminoides | GJ | 茜草科栀子属 Rubiaceae Gardenia | 灌木 Shrub | 园林、食用、药用等 Garden, edible, medicine, etc. |
| 乌冈栎 Quercus phillyraeoides | QP | 壳斗科栎属 Fagaceae Quercus | 乔木 Tree | 木材、酿酒、饲料等 Timber, wine, feed, etc. |
表1 5种耐阴植物的树种属性和用途
Table 1 Species attributes and applications of five shade-enduring plants
| 物种 Species | 缩写 Abbreviation | 科属 Family and genera | 生活型 Life form | 主要用途 Main applications |
|---|---|---|---|---|
| 山杜英 Elaeocarpus sylvestris | ES | 杜英科杜英属 Elaeocarpaceae Elaeocarpus | 乔木 Tree | 园林、木材、防护、药用等 Garden, timber, protection, medicine, etc. |
| 朱砂根 Ardisia crenata | AC | 紫金牛科紫金牛属 Myrsinaceae Ardisia | 灌木 Shrub | 观赏、固土、药用等 Ornamental, soil fixing, medicine, etc. |
| 红茴香 Illicium henryi | IH | 八角科八角属 Illiciaceae Illicium | 乔木 Tree | 观赏、药用等 Ornamental, medicine, etc. |
| 栀子 Gardenia jasminoides | GJ | 茜草科栀子属 Rubiaceae Gardenia | 灌木 Shrub | 园林、食用、药用等 Garden, edible, medicine, etc. |
| 乌冈栎 Quercus phillyraeoides | QP | 壳斗科栎属 Fagaceae Quercus | 乔木 Tree | 木材、酿酒、饲料等 Timber, wine, feed, etc. |
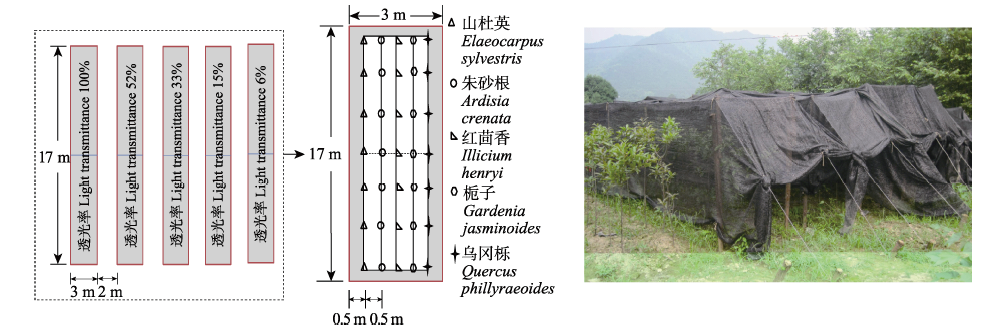
图1 试验设计。
Fig. 1 Experimental design.
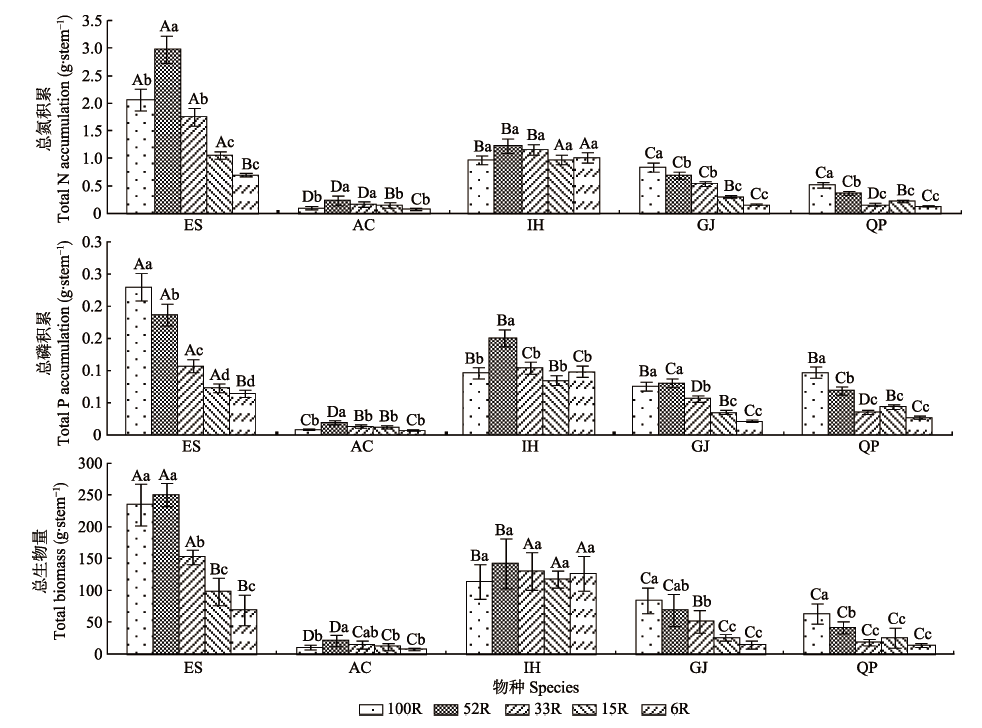
图2 不同光强下5种耐阴植物的总N、P积累和总生物量(平均值±标准偏差)。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。不同小写字母表示同一物种不同光强下总N、P积累和总生物量差异显著(p < 0.05); 不同大写字母表示同一光强下不同物种间总N、P积累和总生物量差异显著(p < 0.05)。
Fig. 2 Total N, P accumulations and biomass of five shade- enduring plants under varying light intensity condition (mean ± SD). AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively. Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels.
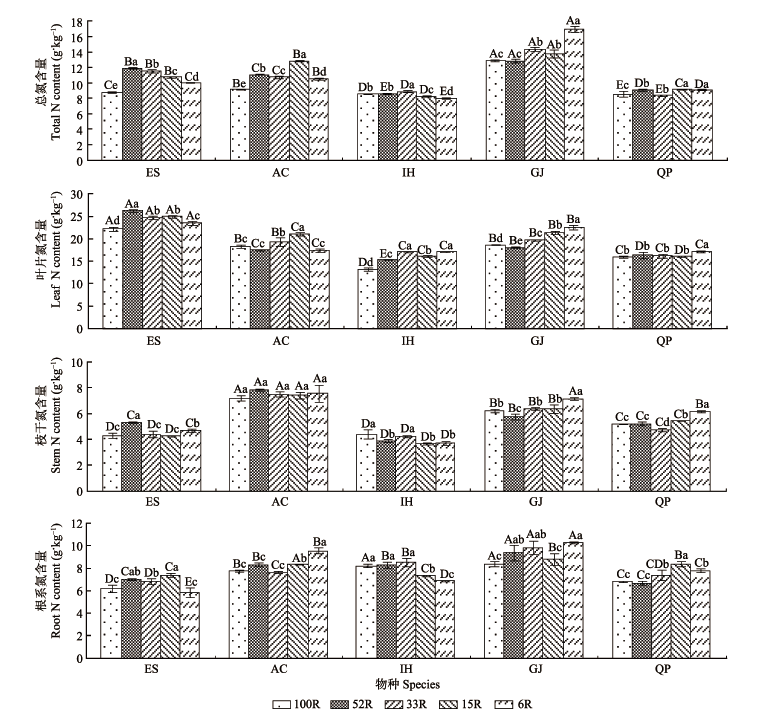
图3 不同光强下5种耐阴植物总N和各器官N含量(平均值±标准偏差)。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。不同小写字母表示同一物种不同光强下N含量差异显著(p < 0.05); 不同大写字母表示同一光强下不同物种间N含量差异显著(p < 0.05)。
Fig. 3 Total N content and organ N contents of five shade-enduring plants under varying light intensity condition (mean ± SD). AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively. Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels.

图4 不同光照强度下植物各器官N、P含量的变异系数。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。
Fig. 4 Coefficients of variation of N and P contents in plant organs under varying light intensity condition. AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides.

图5 不同光强下五种耐阴植物总P和各器官P含量(平均值±标准偏差)。AC, 朱砂根; ES, 山杜英; GJ, 栀子; IH, 红茴香; QP, 乌冈栎。100R、52R、33R、15R和6R分别表示自然光强的100%、52%、33%、15%和6%。不同小写字母表示同一物种不同光强下P含量差异显著(p < 0.05); 不同大写字母表示同一光强下不同物种间P含量差异显著(p < 0.05)。
Fig. 5 Total P content and organ P contents of five shade-enduring plants under varying LIC (mean ± SD). AC, Ardisia crenata; ES, Elaeocarpus sylvestris; GJ, Gardenia jasminoides; IH, Illicium henryi; QP, Quercus phillyraeoides. 100R, 52R, 33R, 15R and 6R represent 100%, 52%, 33%, 15% and 6% of the natural light intensity, respectively. Different lowercase letters indicate significant differences between light intensity conditions of the same species at p < 0.05 levels. Different capital letters indicate significant differences between species of the same light intensity condition at p < 0.05 levels.
| 物种 Species | 相对光强 Relative light intensity | ||||||
|---|---|---|---|---|---|---|---|
| 100R | 52R | 33R | 15R | 6R | |||
| 山杜英 Elaeocarpus sylvestris | 总N:P Total N:P | 9.01 ± 0.59Cd | 15.95 ± 0.53Aa | 16.48 ± 1.19Aa | 14.50 ± 0.29Ab | 10.87 ± 0.38ABc | |
| 叶片N:P Leaf N:P | 15.90 ± 1.19Aab | 18.96 ± 0.59Aab | 19.11 ± 3.11Aa | 17.31 ± 0.45Aab | 15.67 ± 1.79Bb | ||
| 枝干N:P Stem N:P | 6.79 ± 0.60Dc | 15.70 ± 0.02Aa | 16.14 ± 0.40Aa | 12.23 ± 0.37Bb | 6.66 ± 0.10Cc | ||
| 根系N:P Root N:P | 5.17 ± 1.08Db | 8.72 ± 0.59Ba | 9.31 ± 0.83Aa | 9.39 ± 1.34Aa | 8.28 ± 1.06Ba | ||
| 朱砂根 Ardisia crenata | 总N:P Total N:P | 13.57 ± 0.45Aa | 12.62 ± 0.11Bb | 12.26 ± 0.34Bbc | 12.67 ± 0.41Bb | 11.57 ± 0.47Ac | |
| 叶片N:P磷 Leaf N:P | 16.15 ± 0.71Ab | 16.59 ± 0.48Bb | 17.63 ± 2.80Bab | 16.61 ± 1.17Ab | 19.41 ± 0.11Aa | ||
| 枝干N:P Stem N:P | 15.28 ± 1.18Aa | 10.27 ± 0.01Bb | 9.63 ± 0.25Cbc | 8.75 ± 0.44Ccd | 8.28 ± 0.85Bd | ||
| 根系N:P Root N:P | 11.61 ± 0.92Aa | 10.26 ± 0.09Abc | 9.64 ± 0.23Ac | 9.58 ± 0.08Ac | 10.58 ± 0.17Ab | ||
| 红茴香 Illicium henryi | 总N:P Total N:P | 10.14 ± 0.73Ca | 8.16 ± 0.28Da | 11.11 ± 0.04Ca | 11.51 ± 0.07BCa | 10.37 ± 0.02Ba | |
| 叶片N:P Leaf N:P | 10.73 ± 1.06Cb | 9.91 ± 0.28Db | 12.24 ± 0.68Da | 12.09 ± 0.05Ca | 11.87 ± 0.40Ca | ||
| 枝干N:P Stem N:P | 13.22 ± 1.00Bab | 8.16 ± 0.44Cc | 14.05 ± 0.34Bab | 14.53 ± 1.07Aa | 12.37 ± 1.39Ab | ||
| 根系N:P Root N:P | 7.97 ± 0.28Bb | 6.33 ± 0.15Cd | 8.24 ± 0.51Bb | 8.92 ± 0.27Aa | 7.19 ± 0.13Bc | ||
| 栀子 Gardenia jasminoides | 总N:P Total N:P | 12.47 ± 0.24Ba | 10.30 ± 0.15Cb | 11.59 ± 0.91Ca | 9.86 ± 0.42Cb | 11.83 ± 0.29Aa | |
| 叶片N:P Leaf N:P | 15.75 ± 0.51Aa | 14.22 ± 0.34Cb | 15.12 ± 0.91Cab | 15.62 ± 1.26Bab | 16.30 ± 0.48Ba | ||
| 枝干N:P Stem N:P | 10.24 ± 0.72Ca | 6.41 ± 0.39Dc | 8.15 ± 0.55Cb | 5.84 ± 0.06Dcd | 5.14 ± 0.09Dd | ||
| 根系N:P Root N:P | 7.05 ± 0.15Ca | 5.29 ± 0.01Ccd | 5.88 ± 0.89Cbc | 4.75 ± 0.36Bd | 6.48 ± 0.31Cab | ||
| 乌冈栎 Quercus phillyraeoides | 总N:P Total N:P | 5.38 ± 0.01Da | 5.41 ± 0.04Ea | 4.39 ± 0.04Db | 5.23 ± 0.25Da | 4.54 ± 0.05Cb | |
| 叶片N:P Leaf N:P | 11.64 ± 0.23Ba | 9.84 ± 0.15Dbc | 9.24 ± 0.85Ec | 10.39 ± 0.13Db | 9.92 ± 0.16Dbc | ||
| 枝干N:P Stem N:P | 3.65 ± 0.12Ea | 3.32 ± 0.05Eb | 2.98 ± 0.03Dc | 3.57 ± 0.14Ea | 3.59 ± 0.02Ea | ||
| 根系N:P Root N:P | 3.82 ± 0.12Ea | 3.52 ± 0.02Da | 2.79 ± 0.37Db | 3.54 ± 0.35Ca | 2.65 ± 0.02Db | ||
表2 不同光强下5种耐阴植物氮磷比(N:P) (平均值±标准偏差)
Table 2 Nitrogen/phosphorus ratio (N:P) of five shade-enduring plant species under varying light intensity condition (mean ± SD)
| 物种 Species | 相对光强 Relative light intensity | ||||||
|---|---|---|---|---|---|---|---|
| 100R | 52R | 33R | 15R | 6R | |||
| 山杜英 Elaeocarpus sylvestris | 总N:P Total N:P | 9.01 ± 0.59Cd | 15.95 ± 0.53Aa | 16.48 ± 1.19Aa | 14.50 ± 0.29Ab | 10.87 ± 0.38ABc | |
| 叶片N:P Leaf N:P | 15.90 ± 1.19Aab | 18.96 ± 0.59Aab | 19.11 ± 3.11Aa | 17.31 ± 0.45Aab | 15.67 ± 1.79Bb | ||
| 枝干N:P Stem N:P | 6.79 ± 0.60Dc | 15.70 ± 0.02Aa | 16.14 ± 0.40Aa | 12.23 ± 0.37Bb | 6.66 ± 0.10Cc | ||
| 根系N:P Root N:P | 5.17 ± 1.08Db | 8.72 ± 0.59Ba | 9.31 ± 0.83Aa | 9.39 ± 1.34Aa | 8.28 ± 1.06Ba | ||
| 朱砂根 Ardisia crenata | 总N:P Total N:P | 13.57 ± 0.45Aa | 12.62 ± 0.11Bb | 12.26 ± 0.34Bbc | 12.67 ± 0.41Bb | 11.57 ± 0.47Ac | |
| 叶片N:P磷 Leaf N:P | 16.15 ± 0.71Ab | 16.59 ± 0.48Bb | 17.63 ± 2.80Bab | 16.61 ± 1.17Ab | 19.41 ± 0.11Aa | ||
| 枝干N:P Stem N:P | 15.28 ± 1.18Aa | 10.27 ± 0.01Bb | 9.63 ± 0.25Cbc | 8.75 ± 0.44Ccd | 8.28 ± 0.85Bd | ||
| 根系N:P Root N:P | 11.61 ± 0.92Aa | 10.26 ± 0.09Abc | 9.64 ± 0.23Ac | 9.58 ± 0.08Ac | 10.58 ± 0.17Ab | ||
| 红茴香 Illicium henryi | 总N:P Total N:P | 10.14 ± 0.73Ca | 8.16 ± 0.28Da | 11.11 ± 0.04Ca | 11.51 ± 0.07BCa | 10.37 ± 0.02Ba | |
| 叶片N:P Leaf N:P | 10.73 ± 1.06Cb | 9.91 ± 0.28Db | 12.24 ± 0.68Da | 12.09 ± 0.05Ca | 11.87 ± 0.40Ca | ||
| 枝干N:P Stem N:P | 13.22 ± 1.00Bab | 8.16 ± 0.44Cc | 14.05 ± 0.34Bab | 14.53 ± 1.07Aa | 12.37 ± 1.39Ab | ||
| 根系N:P Root N:P | 7.97 ± 0.28Bb | 6.33 ± 0.15Cd | 8.24 ± 0.51Bb | 8.92 ± 0.27Aa | 7.19 ± 0.13Bc | ||
| 栀子 Gardenia jasminoides | 总N:P Total N:P | 12.47 ± 0.24Ba | 10.30 ± 0.15Cb | 11.59 ± 0.91Ca | 9.86 ± 0.42Cb | 11.83 ± 0.29Aa | |
| 叶片N:P Leaf N:P | 15.75 ± 0.51Aa | 14.22 ± 0.34Cb | 15.12 ± 0.91Cab | 15.62 ± 1.26Bab | 16.30 ± 0.48Ba | ||
| 枝干N:P Stem N:P | 10.24 ± 0.72Ca | 6.41 ± 0.39Dc | 8.15 ± 0.55Cb | 5.84 ± 0.06Dcd | 5.14 ± 0.09Dd | ||
| 根系N:P Root N:P | 7.05 ± 0.15Ca | 5.29 ± 0.01Ccd | 5.88 ± 0.89Cbc | 4.75 ± 0.36Bd | 6.48 ± 0.31Cab | ||
| 乌冈栎 Quercus phillyraeoides | 总N:P Total N:P | 5.38 ± 0.01Da | 5.41 ± 0.04Ea | 4.39 ± 0.04Db | 5.23 ± 0.25Da | 4.54 ± 0.05Cb | |
| 叶片N:P Leaf N:P | 11.64 ± 0.23Ba | 9.84 ± 0.15Dbc | 9.24 ± 0.85Ec | 10.39 ± 0.13Db | 9.92 ± 0.16Dbc | ||
| 枝干N:P Stem N:P | 3.65 ± 0.12Ea | 3.32 ± 0.05Eb | 2.98 ± 0.03Dc | 3.57 ± 0.14Ea | 3.59 ± 0.02Ea | ||
| 根系N:P Root N:P | 3.82 ± 0.12Ea | 3.52 ± 0.02Da | 2.79 ± 0.37Db | 3.54 ± 0.35Ca | 2.65 ± 0.02Db | ||
| [1] | Bao SD (2000). Soil Agro-chemistrical Analysis. 3rd edn. Agricultural Press of China, Beijing. 42. (in Chinese)[鲍士旦 (2000). 土壤农化分析. 第三版.中国农业出版社, 北京. 42.] |
| [2] | Cheng XR, Zhou JH, Chen YH, Wu KZ, Yu MK (2016). Effects of light intensity on phenotypic plasticity ofGardenia jasminoides seedlings. Acta Agriculturae Universitatis Jiangxiensis, 38, 180-186. (in Chinese with English abstract)[成向荣, 周俊宏, 陈永辉, 武克壮, 虞木奎 (2016). 山栀子幼苗表型可塑性对不同光环境的响应. 江西农业大学学报, 38, 180-186.] |
| [3] | Donovan LA, Maherali H, Caruso CM, Huber H, Kroon HD (2011). The evolution of the worldwide leaf economics spectrum.Trends in Ecology & Evolution, 26, 88-95. |
| [4] | Groot DC, Boogaard R, Marcelis LM, Harbinson J, Lambers H (2003). Contrasting effects of N and P deprivation on the regulation of photosynthesis in tomato plants in relation to feedback limitation.Journal of Experimental Botany, 54, 1957-1967. |
| [5] | Guan M, Jin ZX, Wang Q, Li YL, Zuo W (2012). Response of photosynthesis traits of dominant plant species to different light regimes in the secondary forest in the area of Qiandao Lake, Zhejiang, China.Chinese Journal of Applied Ecology, 25, 1615-1622. (in Chinese with English abstract)[管铭, 金则新, 王强, 李月灵, 左威 (2012). 千岛湖次生林优势种植物光合特性对不同光环境的响应. 应用生态学报, 25, 1615-1622.] |
| [6] | Guan YX, Lin B, Ling BY (2000). Interaction effects of light density and nitrogen on maize leaf pigments, chlorophyll fluorescence and energy transition.Plant Nutrition and Fertilizer Science, 6, 152-158. (in Chinese with English abstract)[关义新, 林葆, 凌碧莹 (2000). 光氮互作对玉米叶片光合色素及其荧光特性与能量转换的影响. 植物营养与肥料学报, 6, 152-158.] |
| [7] | Güsewell S (2004). N:P ratios in terrestrial plants: Variation and functional significance.New Phytologist, 164, 243-266. |
| [8] | Ingestad T, Ågren GI (1988). Nutrient uptake and allocation at steady-state nutrition.Physiologia Plantarum, 72, 450-459. |
| [9] | Kang M, Xie YM, Xu Y, Xu YL, Yan ER (2013). Within- community variability of plant leaf N and P contents in Tiantong, Zhejiang Province.Journal of East China Normal University (Natural Science), (2), 20-29. (in Chinese with English abstract)[康蒙, 谢一鸣, 许月, 徐艺露, 阎恩荣 (2013). 浙江天童植物叶片氮磷含量的群落内变异. 华东师范大学学报(自然科学版), (2), 20-29.] |
| [10] | Koerselman W, Meuleman AFM (1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation.Journal of Applied Ecology, 33, 1441-1450. |
| [11] | Li DF, Yu SL, Wang GX, Fang WW (2015). Environmental heterogeneity and mechanism of stoichiometry properties of vegetative organs in dominant shrub communities across the Loess Plateau.Chinese Journal of Plant Ecology, 39, 453-465. (in Chinese with English abstract)[李单凤, 于顺利, 王国勋, 方伟伟 (2015). 黄土高原优势灌丛营养器官化学计量特征的环境分异和机制. 植物生态学报, 39, 453-465.] |
| [12] | Li YL, Mao W, Zhao XY, Zhang TH (2010). Leaf nitrogen and phosphorus stoichiometry in typical desert and desertified regions, North China.Environmental Science, 31, 1716-1725. (in Chinese with English abstract)[李玉霖, 毛伟, 赵学勇, 张铜会 (2010). 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究. 环境科学, 31, 1716-1725.] |
| [13] | Li ZA, Wang BS, Kong GH, Zhang ZP, Weng H (1999). The element content of plant inCryptocarya concinna community in Dinghushan evergreen monsoon broad-leaf forest. Acta Phytoecologica Sinica, 23, 411-417. (in Chinese with English abstract)[李志安, 王伯荪, 孔国辉, 张祝平, 翁轰 (1999). 鼎湖山季风常绿阔叶林黄果厚壳桂群落植物元素含量特征. 植物生态学报, 23, 411-417.] |
| [14] | Liu C, Wang Y, Wang N, Wang GX (2012). Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial ecosystems: A review.Chinese Journal of Plant Ecology, 36, 1205-1216. (in Chinese with English abstract)[刘超, 王洋, 王楠, 王根轩 (2012). 陆地生态系统植被氮磷化学计量研究进展. 植物生态学报, 36, 1205-1216.] |
| [15] | Liu GS, Yun F, Shi HZ, Wang K, Zhang CH, Song J (2010). Effects of cooperation of light and nitrogen on the content of nitric compound, antioxidant system and quality of flue- cured tobacco.Scientia Agricultura Sinica, 43, 3732-3741. (in Chinese with English abstract)[刘国顺, 云菲, 史宏志, 王可, 张春华, 宋晶 (2010). 光、氮及其互作对烤烟含氮化合物含量、抗氧化系统及品质的影响. 中国农业科学, 43, 3732-3741.] |
| [16] | Liu WF, Fan HB (2008). Characteristics of nitrogen cycles in the ecosystem of mixed stands of Masson pine and broadleaved forests.Journal of Beijing Forestry University, 30(6), 52-58. (in Chinese with English abstract)[刘文飞, 樊后保 (2008). 马尾松阔叶树混交林生态系统的氮素循环特征. 北京林业大学学报, 30(6), 52-58.] |
| [17] | Liu XZ, Kang SZ (2002). Effects of shading on photosynthesis, dry matter partitioning and N, P, K concentrations in leaves of tomato plants at different growth stages.Acta Ecologica Sinica, 22, 2264-2271. (in Chinese with English abstract)[刘贤赵, 康绍忠 (2002). 不同生长阶段遮荫对番茄光合作用、干物质分配与叶N、P、K的影响. 生态学报, 22, 2264-2271.] |
| [18] | Matzek V, Vitousek PM (2009). N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis.Ecology Letters, 12, 765-771. |
| [19] | Qiao XR (2007). Effects of Light Intensity on Growth Photosynthetic Characteristics and Quality of Flue-cured Tobacco. Master degree dissertation, Henan Agricultural University, Zhengzhou. (in Chinese with English abstract)[乔新荣 (2007). 光照强度对烤烟生长发育、光合特性及品质的影响. 硕士学位论文, 河南农业大学, 郑州.] |
| [20] | Shi JG, Zhu KL, Cao HY, Dong ST, Liu P, Zhao B, Zhang JW (2015). Effect of light from flowering to maturity stage on dry matter accumulation and nutrient absorption of summer maize.Chinese Journal of Applied Ecology, 26, 46-52.(in Chinese with English abstract). [史建国, 朱昆仑, 曹慧英, 董树亭, 刘鹏, 赵斌, 张吉旺 (2015). 花粒期光照对夏玉米干物质积累和养分吸收的影响. 应用生态学报, 26, 46-52.] |
| [21] | Shi JZ, Wang TD (1994). A mechanistic model describing the photosynthate partitioning during vegetative phase.Acta Botanica Sinica, 36, 181-189. (in Chinese with English abstract)[施建忠, 王天铎 (1994). 植物营养生长期同化物分配的机理模型. 植物学报, 36, 181-189.] |
| [22] | Sims L, Pastor J, Lee T, Dewey B (2012). Nitrogen, phosphorus and light effects on growth and allocation of biomass and nutrients in wild rice.Oecologia, 170, 65-76. |
| [23] | Suriyagoda LB, Lambers H, Renton M, Renton M, Ryan MH (2012). Growth, carboxylate exudates and nutrient dynamics in three herbaceous perennial plant species under low, moderate and high phosphorus supply.Plant and Soil, 358, 105-117. |
| [24] | Thompson K, Parkinson JA, Band SR, Spencer RE (1997). A comparative study of leaf nutrient concentrations in a regional herbaceous flora.New Phytologist, 136, 679-689. |
| [25] | Valladares F, Martinez FE, Balaguer L, Corona EP, Manrique E (2000). Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource- use strategy?New Phytologist, 148, 79-91. |
| [26] | Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010). Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions.Ecological Applications, 20, 5-15. |
| [27] | Wang H, Cai ZQ, Cai CT, Zhang SX (2008). A comparative study of two tropical pioneer species with different life span under different light and nutrient conditions.Journal of Wuhan Botanical Research, 26, 134-141. (in Chinese with English abstract)[王辉, 蔡志全, 蔡传涛, 张硕新 (2008). 不同光照和营养条件下两种不同寿命热带先锋种的对比研究. 武汉植物学研究, 26, 134-141.] |
| [28] | Wang SQ, Yu GR (2008). Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus element.Acta Ecologica Sinica, 28, 3937-3947. (in Chinese with English abstract)[王绍强, 于贵瑞 (2008). 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 28, 3937-3947.] |
| [29] | Wang ZN, Yang HM (2013). Response of ecological stoichiometry of carbon, nitrogen and phosphorus in plants to abiotic environmental factors.Pratacultural Science, 30, 927-934. (in Chinese with English abstract)[王振南, 杨惠敏 (2013). 植物碳氮磷生态化学计量对非生物因子的响应. 草业科学, 30, 927-934.] |
| [30] | Xu QX (2013). Effect of Thinning on Soil Physicochemical Property and Parbon Ptorage of the Patural Larix gmelinii Forest in Great Xing’an Mountains. Master degree dissertation, Northeast Forestry University, Harbin. (in Chinese with English abstract)[徐庆祥 (2013). 抚育间伐对大兴安岭兴安落叶松天然林碳储量的影响. 硕士学位论文, 东北林业大学, 哈尔滨.] |
| [31] | Xu XY, Xiao L, Wang MH, Zhang HX (2015). A comprehensive evaluation system for anti-typhoon performance of trees in coastal areas.Journal of Zhejiang A & F University, 32, 516-522. (in Chinese with English abstract)[许秀玉, 肖莉, 王明怀, 张华新 (2015). 沿海抗台风树种评价体系构建与选择. 浙江农林大学学报, 32, 516-522.] |
| [32] | Yan YF, Fang SZ, Tian Y, Song H, Tun XJ (2014). The response of understory plant diversity and nutrient accumulation to stand structure of poplar plantation.Chinese Journal of Ecology, 33, 1170-1177. (in Chinese with English abstract)[燕亚飞, 方升佐, 田野, 宋浩, 囤兴建 (2014). 林下植物多样性及养分积累量对杨树林分结构的响应. 生态学杂志, 33, 1170-1177.] |
| [33] | Yang X, Tang ZY, Ji C, Liu HY, Ma WH, Mohhamot A, Shi ZY, Sun W, Wang T, Wang XP, Wu X, Yu SL, Yue M, Zheng CY (2014). Scaling of nitrogen and phosphorus across plant organs in shrub land biomes across Northern China.Scientific Reports, 4, 5448. doi: 10.1038/srep05448. |
| [34] | Yin HJ, Lai T, Cheng XY, Jiang XM, Liu Q (2008). Warming effects on growth and physiology of seedlings of Betula albo-sinensis and Abies faxoniana under two contrasting light conditions in subalpine coniferous forest of western Sichuan, China. Journal of Plant Ecology (Chinese Version), 32, 1072-1083. (in Chinese with English abstract)[尹华军, 赖挺, 程新颖, 蒋先敏, 刘庆 (2008). 增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响. 植物生态学报, 32, 1072-1083.] |
| [35] | Zhao P, Kriebitzseh W, Zhang ZQ (1999). Gas exchange, chlorophyll and nitrogen contents in leaves of three common trees in middle Europe under two contrasting light regime.Journal of Tropical and Subtropical Botany, 7, 133-139. (in Chinese with English abstract)[赵平, Kriebitzseh W, 张志权 (1999). 欧洲3种常见乔木幼苗在两种光环境下叶片的气体交换、叶绿素含量和氮素含量. 热带亚热带植物学报, 7, 133-139.] |
| [36] | Zheng SX, Shangguan ZP (2007). Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China.Trees, 21, 357-370. |
| [1] | 梁逸娴, 王传宽, 臧妙涵, 上官虹玉, 刘逸潇, 全先奎. 落叶松径向生长和生物量分配对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 459-468. |
| [2] | 耿雪琪, 唐亚坤, 王丽娜, 邓旭, 张泽凌, 周莹. 氮添加增加中国陆生植物生物量并降低其氮利用效率[J]. 植物生态学报, 2024, 48(2): 147-157. |
| [3] | 吴晨, 陈心怡, 刘源豪, 黄锦学, 熊德成. 增温对森林细根生长、死亡及周转特征影响的研究进展[J]. 植物生态学报, 2023, 47(8): 1043-1054. |
| [4] | 陈雪纯, 刘虹, 朱少琦, 孙铭遥, 宇振荣, 王庆刚. 漓江流域不同弃耕年限下4种常见草本植物功能性状种内变化及其影响因素[J]. 植物生态学报, 2023, 47(4): 559-570. |
| [5] | 陈心怡, 吴晨, 黄锦学, 熊德成. 增温对林木细根物候影响的研究进展[J]. 植物生态学报, 2023, 47(11): 1471-1482. |
| [6] | 刘艳杰, 刘玉龙, 王传宽, 王兴昌. 东北温带森林5个羽状复叶树种叶成本-效益关系比较[J]. 植物生态学报, 2023, 47(11): 1540-1550. |
| [7] | 刘洋, 马煦, 邸楠, 曾子航, 付海曼, 李新, 席本野. 毛白杨根系液流与水力再分配特征[J]. 植物生态学报, 2023, 47(1): 123-133. |
| [8] | 姚萌, 康荣华, 王盎, 马方园, 李靳, 台子晗, 方运霆. 利用15N示踪技术研究木荷与马尾松幼苗叶片对NO2的吸收与分配[J]. 植物生态学报, 2023, 47(1): 114-122. |
| [9] | 韩广轩, 王法明, 马俊, 肖雷雷, 初小静, 赵明亮. 滨海盐沼湿地蓝色碳汇功能、形成机制及其增汇潜力[J]. 植物生态学报, 2022, 46(4): 373-382. |
| [10] | 罗源林, 马文红, 张芯毓, 苏闯, 史亚博, 赵利清. 内蒙古锦鸡儿属植物地理替代分布种的功能性状沿环境梯度的变化[J]. 植物生态学报, 2022, 46(11): 1364-1375. |
| [11] | 林夏珍, 刘林, 董婷婷, 方琦博, 郭庆学. 非结构性碳水化合物与氮分配对美洲黑杨和青杨耐盐能力的影响[J]. 植物生态学报, 2021, 45(9): 961-971. |
| [12] | 靳川, 李鑫豪, 蒋燕, 徐铭泽, 田赟, 刘鹏, 贾昕, 查天山. 黑沙蒿光合能量分配组分在生长季的相对变化与调控机制[J]. 植物生态学报, 2021, 45(8): 870-879. |
| [13] | 武运涛, 杨森, 王欣, 黄俊胜, 王斌, 刘卫星, 刘玲莉. 草地土壤有机质不同组分氮库对长期氮添加的响应[J]. 植物生态学报, 2021, 45(7): 790-798. |
| [14] | 韩广轩, 李隽永, 屈文笛. 氮输入对滨海盐沼湿地碳循环关键过程的影响及机制[J]. 植物生态学报, 2021, 45(4): 321-333. |
| [15] | 丁键浠, 周蕾, 王永琳, 庄杰, 陈集景, 周稳, 赵宁, 宋珺, 迟永刚. 叶绿素荧光主动与被动联合观测应用前景[J]. 植物生态学报, 2021, 45(2): 105-118. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19