
植物生态学报 ›› 2022, Vol. 46 ›› Issue (11): 1364-1375.DOI: 10.17521/cjpe.2021.0491
所属专题: 植物功能性状
罗源林1, 马文红1,2,*( ), 张芯毓1, 苏闯1, 史亚博1, 赵利清1,2
), 张芯毓1, 苏闯1, 史亚博1, 赵利清1,2
收稿日期:2021-12-27
接受日期:2022-06-23
出版日期:2022-11-20
发布日期:2022-07-06
通讯作者:
*马文红(whma@imu.edu.cn)
基金资助:
LUO Yuan-Lin1, MA Wen-Hong1,2,*(), ZHANG Xin-Yu1, SU Chuang1, SHI Ya-Bo1, ZHAO Li-Qing1,2
Received:2021-12-27
Accepted:2022-06-23
Online:2022-11-20
Published:2022-07-06
Contact:
*MA Wen-Hong(whma@imu.edu.cn)
Supported by:摘要:
锦鸡儿属(Caragana)植物是蒙古高原干旱生态系统的重要组成成分, 沿着环境梯度形成了极为明显的地理替代分布。为了检验这些替代分布种对气候和土壤环境变化的适应性, 该研究在内蒙古地区选择了41个样地、9种锦鸡儿属植物, 测定了这些植物的株高(h)、叶性状(叶干物质含量(LDMC)、比叶面积(SLA)、叶面积(LA)、叶片氮含量(LNC)、叶片磷含量(LPC)和氮磷比(N:P))和茎性状(茎组织密度(STD)), 分析了不同功能性状之间及功能性状与气候和土壤特征之间的关系。结果表明: 9种植物的功能性状沿着气候梯度呈现出明显的变化趋势, 干燥度指数解释了大部分功能性状29%以上的变异(N:P除外), 随着干旱程度增强, 植物的h、SLA、LA、LNC和LPC降低, 而LDMC和STD增加。土壤氮含量也在一定程度上影响植物的功能性状, 尤其是反映植物竞争光和空间等资源的重要性状(h)以及反映植物生长速率的关键性状(SLA、LNC和STD), 都与土壤氮供应能力显著相关。而土壤pH对锦鸡儿属植物的性状解释度较低。整体上, 在属水平上植物功能性状之间具有较强的共变关系, 但在种水平上, 分布最广的柠条锦鸡儿(C. korshinskii)和小叶锦鸡儿(C. microphylla)具有较强的性状共变关系, 其他分布范围较小物种功能性状间的共变关系相对较弱。这一结果意味着植物功能性状在属内的变化对气候和土壤环境变化产生一定的适应, 有助于预测全球变化背景下群落结构以及功能多样性的变化。分布范围较小的植物种内功能性状间的共变关系微弱, 意味着植物经济谱理论不一定适合在局域尺度上解释植物在个体水平上对资源的利用和对环境变化的适应策略。
罗源林, 马文红, 张芯毓, 苏闯, 史亚博, 赵利清. 内蒙古锦鸡儿属植物地理替代分布种的功能性状沿环境梯度的变化. 植物生态学报, 2022, 46(11): 1364-1375. DOI: 10.17521/cjpe.2021.0491
LUO Yuan-Lin, MA Wen-Hong, ZHANG Xin-Yu, SU Chuang, SHI Ya-Bo, ZHAO Li-Qing. Variation of functional traits of alternative distribution of Caragana species along environmental gradients in Nei Mongol, China. Chinese Journal of Plant Ecology, 2022, 46(11): 1364-1375. DOI: 10.17521/cjpe.2021.0491
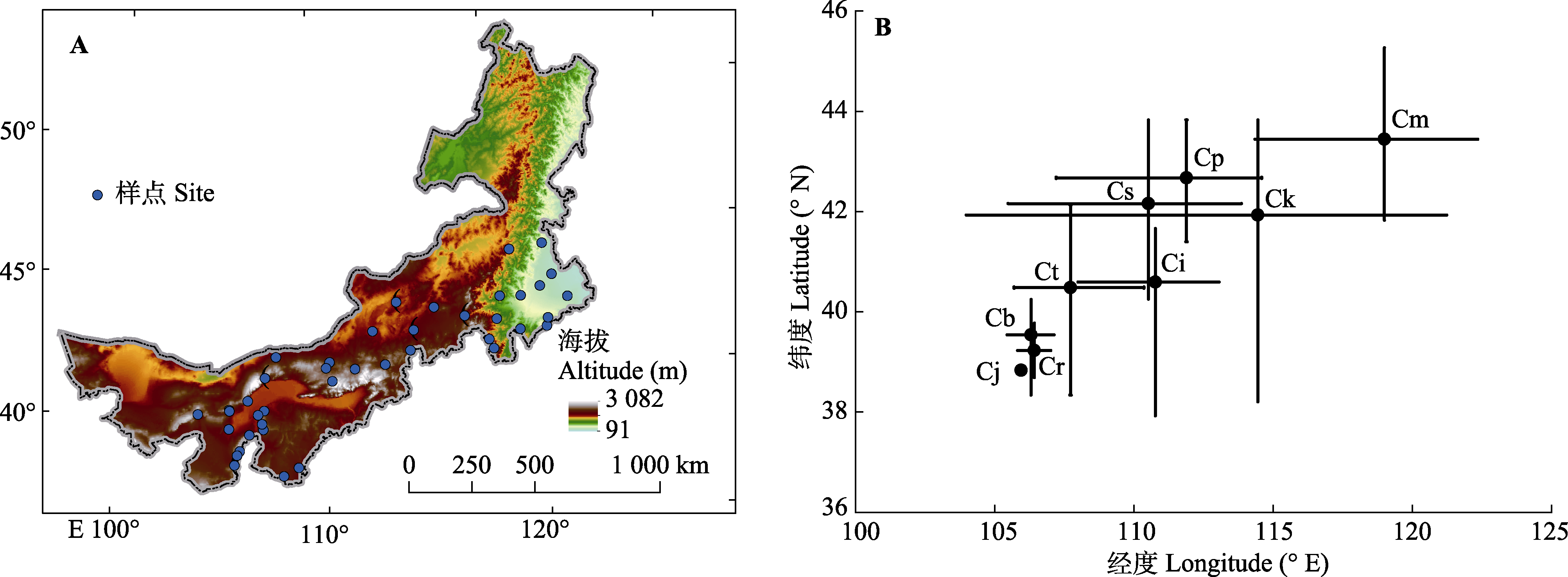
图1 内蒙古锦鸡儿属植物样点分布图(A)和取样地理范围(B)。B中黑色圆点代表经纬度平均值。Cb, 矮脚锦鸡儿; Ci, 中间锦鸡儿; Cj, 鬼箭锦鸡儿; Ck, 柠条锦鸡儿; Cm, 小叶锦鸡儿; Cp, 矮锦鸡儿; Cr, 荒漠锦鸡儿; Cs, 狭叶锦鸡儿; Ct, 毛刺锦鸡儿。
Fig. 1 Distribution map of observation sites (A) and geographical range of observation sites (B) of Caragana species in Nei Mongol. Black solid points in B represent the average latitude and longitude of the site locations of different species of Caragana species. Cb, C. brachypoda; Ci, C. intermedia; Cj, C. jubata; Ck, C. korshinskii; Cm, C. microphylla; Cp, C. pygmaea; Cr, C. roborovskyi; Cs, C. stenophylla; Ct, C. tibetica.
| 物种 Species | 干燥度指数范围 Aridity index range | 土壤氮含量范围 Soil nitrogen content range (mg·g-1) | 土壤pH范围 Soil pH range | 样点数 Sample number |
|---|---|---|---|---|
| 矮脚锦鸡儿 C. brachypoda | 0.17-0.33 | 0.2-0.5 | 8.49-8.90 | 5 |
| 毛刺锦鸡儿 C. tibetica | 0.25-0.43 | 0.3-3.2 | 8.55-8.86 | 4 |
| 荒漠锦鸡儿 C. roborovskyi | 0.31-0.45 | 0.7-3.7 | 8.07-8.78 | 2 |
| 矮锦鸡儿 C. pygmaea | 0.30-0.49 | 0.3-0.3 | 8.69-8.87 | 3 |
| 柠条锦鸡儿 C. korshinskii | 0.15-0.75 | 0.2-1.3 | 8.01-9.12 | 12 |
| 狭叶锦鸡儿 C. stenophylla | 0.22-0.69 | 0.5-1.2 | 8.54-8.57 | 4 |
| 中间锦鸡儿 C. intermedia | 0.30-0.59 | 0.3-3.0 | 8.62-8.87 | 5 |
| 小叶锦鸡儿 C. microphylla | 0.52-0.75 | 0.3-4.4 | 6.39-8.57 | 10 |
| 鬼箭锦鸡儿 C. jubata | 0.88 | 6.3 | 7.17 | 1 |
表1 内蒙古锦鸡儿属物种列表
Table 1 Caragana species in Nei Mongol, China
| 物种 Species | 干燥度指数范围 Aridity index range | 土壤氮含量范围 Soil nitrogen content range (mg·g-1) | 土壤pH范围 Soil pH range | 样点数 Sample number |
|---|---|---|---|---|
| 矮脚锦鸡儿 C. brachypoda | 0.17-0.33 | 0.2-0.5 | 8.49-8.90 | 5 |
| 毛刺锦鸡儿 C. tibetica | 0.25-0.43 | 0.3-3.2 | 8.55-8.86 | 4 |
| 荒漠锦鸡儿 C. roborovskyi | 0.31-0.45 | 0.7-3.7 | 8.07-8.78 | 2 |
| 矮锦鸡儿 C. pygmaea | 0.30-0.49 | 0.3-0.3 | 8.69-8.87 | 3 |
| 柠条锦鸡儿 C. korshinskii | 0.15-0.75 | 0.2-1.3 | 8.01-9.12 | 12 |
| 狭叶锦鸡儿 C. stenophylla | 0.22-0.69 | 0.5-1.2 | 8.54-8.57 | 4 |
| 中间锦鸡儿 C. intermedia | 0.30-0.59 | 0.3-3.0 | 8.62-8.87 | 5 |
| 小叶锦鸡儿 C. microphylla | 0.52-0.75 | 0.3-4.4 | 6.39-8.57 | 10 |
| 鬼箭锦鸡儿 C. jubata | 0.88 | 6.3 | 7.17 | 1 |
| 变异来源 Source of variation | h | LDMC | SLA | STD | LA | LNC | LPC | N:P | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | |
| AI | 24.70 | 31.06*** | 32.72 | 40.80*** | 21.57 | 33.68*** | 16.61 | 29.76*** | 87.13 | 67.39*** | 20.45 | 31.80*** | 16.69 | 34.89*** | 2.77 | 8.42 |
| SN | 15.70 | 19.74*** | 6.99 | 8.71* | 5.61 | 8.76* | 8.41 | 15.07** | 0.26 | 0.20 | 14.93 | 23.21*** | 0.82 | 1.72 | 1.17 | 3.54 |
| SpH | 0.00 | 0.00 | 2.26 | 2.81 | 0.26 | 0.40 | 0.87 | 1.56 | 0.20 | 0.15 | 0.32 | 0.50 | 0.24 | 0.51 | 0.00 | 0.00 |
| AI × SN | 1.10 | 1.38 | 2.21 | 2.76 | 0.38 | 0.60 | 0.32 | 0.57 | 7.74 | 5.99** | 2.49 | 3.87 | 0.63 | 1.32 | 0.03 | 0.08 |
| AI × SpH | 1.17 | 1.47 | 1.38 | 1.72 | 2.52 | 3.94 | 0.04 | 0.07 | 0.79 | 0.61 | 0.07 | 0.11 | 1.18 | 2.48 | 3.25 | 9.85 |
| SN × SpH | 0.28 | 0.35 | 0.01 | 0.02 | 0.18 | 0.28 | 0.39 | 0.70 | 0.60 | 0.46 | 0.25 | 0.39 | 0.17 | 0.35 | 0.01 | 0.03 |
| AI × SN × SpH | 4.59 | 5.77* | 2.62 | 3.27 | 1.54 | 2.40 | 0.18 | 0.32 | 0.58 | 0.45 | 0.81 | 1.25 | 0.09 | 0.20 | 0.73 | 2.20 |
表2 内蒙古锦鸡儿属植物功能性状变异来源
Table 2 Source of variation in plant function traits for Caragana species in Nei Mongol, China
| 变异来源 Source of variation | h | LDMC | SLA | STD | LA | LNC | LPC | N:P | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | F | SS% | |
| AI | 24.70 | 31.06*** | 32.72 | 40.80*** | 21.57 | 33.68*** | 16.61 | 29.76*** | 87.13 | 67.39*** | 20.45 | 31.80*** | 16.69 | 34.89*** | 2.77 | 8.42 |
| SN | 15.70 | 19.74*** | 6.99 | 8.71* | 5.61 | 8.76* | 8.41 | 15.07** | 0.26 | 0.20 | 14.93 | 23.21*** | 0.82 | 1.72 | 1.17 | 3.54 |
| SpH | 0.00 | 0.00 | 2.26 | 2.81 | 0.26 | 0.40 | 0.87 | 1.56 | 0.20 | 0.15 | 0.32 | 0.50 | 0.24 | 0.51 | 0.00 | 0.00 |
| AI × SN | 1.10 | 1.38 | 2.21 | 2.76 | 0.38 | 0.60 | 0.32 | 0.57 | 7.74 | 5.99** | 2.49 | 3.87 | 0.63 | 1.32 | 0.03 | 0.08 |
| AI × SpH | 1.17 | 1.47 | 1.38 | 1.72 | 2.52 | 3.94 | 0.04 | 0.07 | 0.79 | 0.61 | 0.07 | 0.11 | 1.18 | 2.48 | 3.25 | 9.85 |
| SN × SpH | 0.28 | 0.35 | 0.01 | 0.02 | 0.18 | 0.28 | 0.39 | 0.70 | 0.60 | 0.46 | 0.25 | 0.39 | 0.17 | 0.35 | 0.01 | 0.03 |
| AI × SN × SpH | 4.59 | 5.77* | 2.62 | 3.27 | 1.54 | 2.40 | 0.18 | 0.32 | 0.58 | 0.45 | 0.81 | 1.25 | 0.09 | 0.20 | 0.73 | 2.20 |
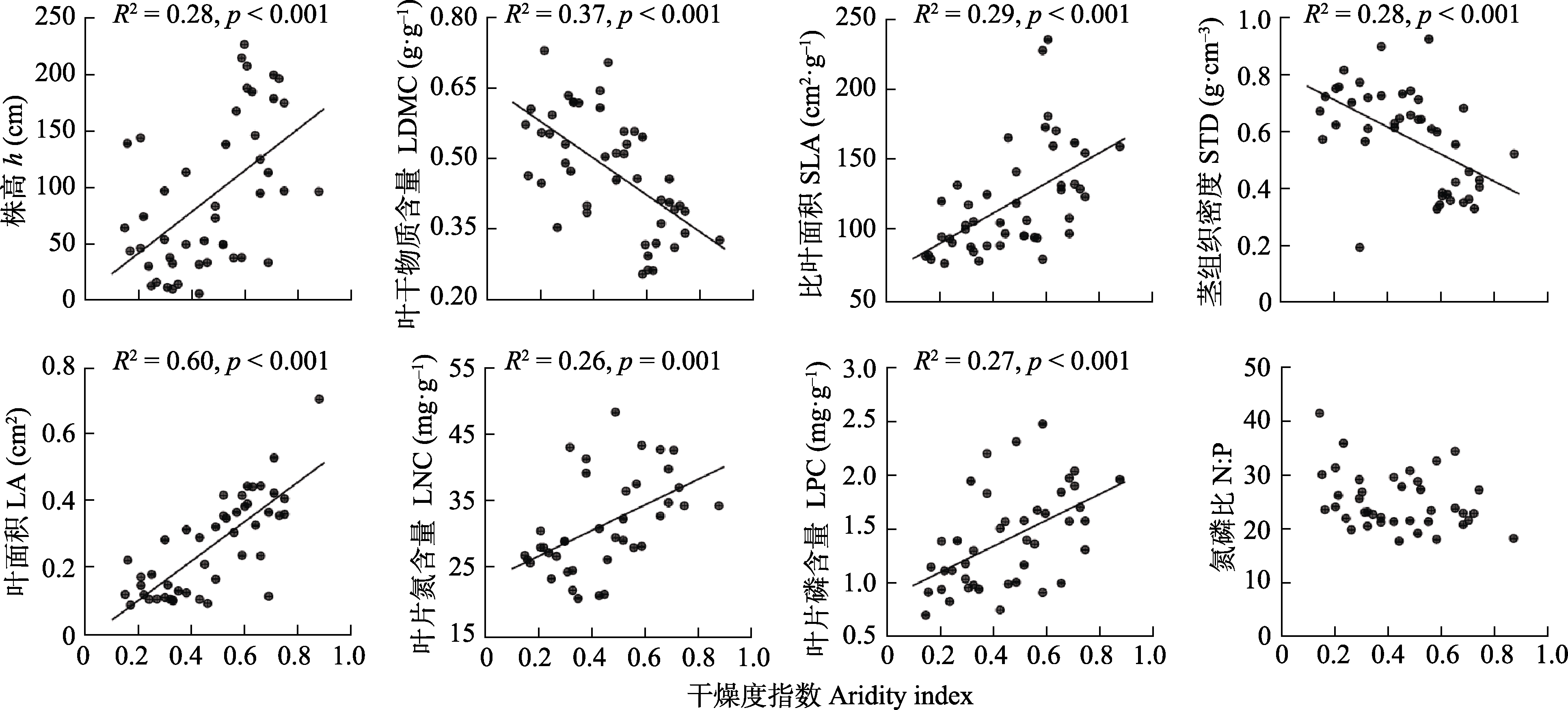
图2 内蒙古锦鸡儿属植物功能性状与干燥度指数的关系。
Fig. 2 Relationships between functional traits of Caragana species and aridity index in Nei Mongol, China. h, height; LA, leaf area; LDMC, leaf dry matter content; LNC, leaf nitrogen content; LPC, leaf phosphorus content; N:P, leaf nitrogen content to phosphorus content ratio; SLA, specific leaf area; STD, stem tissue density.
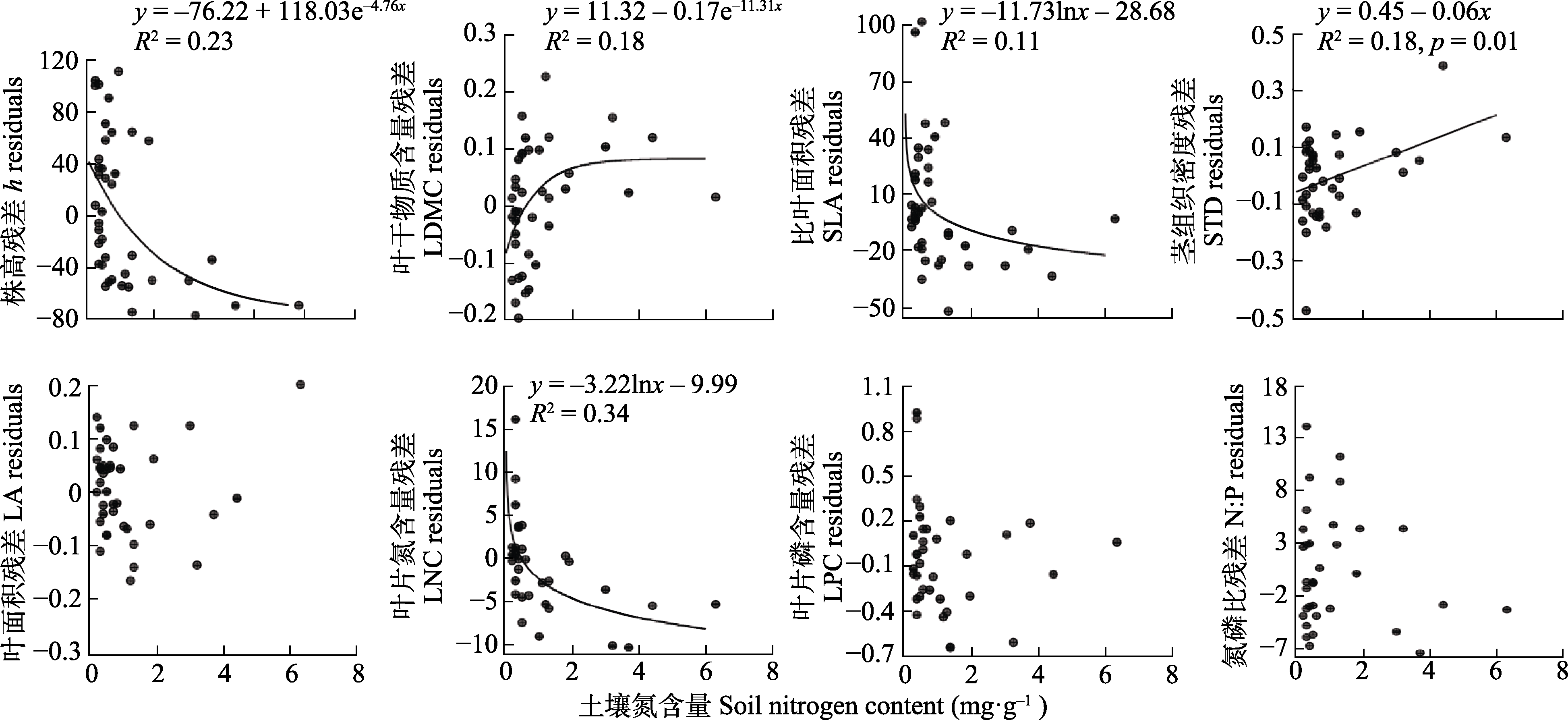
图3 内蒙古锦鸡儿属植物功能性状与土壤氮含量的关系。
Fig. 3 Relationships between functional traits of Caragana species and soil nitrogen content in Nei Mongol. h, height; LA, leaf area; LDMC, leaf dry matter content; LNC, leaf nitrogen content; LPC, leaf phosphorus content; N:P, leaf nitrogen content to phosphorus content ratio; SLA, specific leaf area; STD, stem tissue density.
| x | y | 锦鸡儿属 C. species | 矮锦鸡儿 C. pygmaea | 矮脚锦鸡儿 C. brachypoda | 鬼箭锦鸡儿 C. jubata | 荒漠锦鸡儿 C. roborovskyi | 毛刺锦鸡儿 C. tibetica | 柠条锦鸡儿 C. korshinskii | 狭叶锦鸡儿 C. stenophylla | 小叶锦鸡儿 C. microphylla | 中间锦鸡儿 C. intermedia | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | ||
| h | LDMC | -0.31*** | 0.21 | 0.61 | -1.40 | 0.50* | -1.03 | -0.39 | 0.29 | -0.15** | -0.04 | -0.39 | 0.16 | -0.56*** | 0.80 | 0.86 | -1.66 | -0.45*** | 0.52 | -0.26 | 0.19 |
| SLA | 0.33*** | 1.45 | 0.75 | 0.70 | -0.47 | 2.69 | 0.70 | 0.81 | -0.18* | 2.27 | -0.51 | 2.43 | 0.69*** | 0.62 | -0.79 | 3.24 | 0.54*** | 1.00 | 0.23** | 1.56 | |
| STD | -0.43*** | 0.52 | -1.40 | 2.33 | 0.43 | -0.76 | -0.49 | 0.69 | 0.21 | -0.55 | -0.23 | -0.02 | -0.75** | 1.29 | 0.95 | -1.74 | -0.65*** | 0.97 | -0.83 | 1.16 | |
| LA | 0.64*** | -1.83 | 1.02 | -2.71 | 0.51 | -1.75 | -0.89 | 1.60 | 0.24* | -1.10 | 0.64* | -1.53 | 0.98*** | -2.64 | 0.42 | -1.66 | 0.38** | -1.21 | 0.47 | -1.36 | |
| LNC | 0.25*** | 1.06 | 0.68 | 0.37 | 0.19 | 1.15 | 0.38 | 0.77 | -0.10** | 1.49 | 0.17 | 1.16 | 0.42* | 0.65 | -0.54 | 2.41 | 0.26*** | 1.04 | 0.20 | 1.12 | |
| LPC | 0.37*** | -0.55 | 1.14 | -1.76 | -0.46 | 0.72 | 0.79 | -1.27 | 0.51 | -0.65 | 0.41** | -0.44 | 0.88*** | -1.74 | -0.80 | 1.45 | 0.36*** | -0.55 | 0.44 | -0.71 | |
| N:P | -0.26 | 1.82 | -0.75 | 2.65 | 0.45 | 0.72 | -0.60 | 2.44 | -0.55 | 2.06 | -0.33** | 1.69 | -0.58*** | 2.64 | 0.31 | 0.87 | -0.23 | 1.84 | 0.38 | 0.74 | |
| LDMC | SLA | -1.07*** | 1.67 | -1.24 | 1.58 | -0.95*** | 1.71 | -1.79 | 1.33 | 1.21 | 2.32 | -1.28 | 1.67 | -1.23*** | 1.60 | -0.92 | 1.71 | -1.20*** | 1.62 | -0.87 | 1.72 |
| STD | 1.32*** | 0.20 | 2.23 | 0.64 | -0.85 | -0.41 | -1.26 | -0.90 | -0.75 | -0.41 | -0.49 | -0.31 | 1.33*** | 0.22 | 1.11 | 0.11 | 1.35*** | 0.20 | 3.19 | 0.55 | |
| LA | -2.07*** | -1.39 | 1.69 | -0.34 | -1.03 | -1.32 | -2.27 | -1.26 | -1.61* | -1.16 | -1.51 | -1.24 | -1.74*** | -1.26 | 0.49 | -0.85 | -0.83* | -0.78 | 1.79 | -0.06 | |
| LNC | -0.86*** | 1.21 | -1.13 | 1.18 | -0.37 | 1.31 | -0.98 | 1.05 | 0.66** | 1.51 | -0.43 | 1.24 | -0.74*** | 1.23 | -0.64*** | 1.35 | -0.75*** | 1.27 | -0.77 | 1.27 | |
| LPC | -1.32*** | -0.33 | -1.88** | -0.43 | -0.91* | -0.22 | -2.01 | -0.69 | -3.36 | -0.78 | -1.04 | -0.27 | -1.69*** | -0.55 | -0.94*** | -0.11 | -1.06** | -0.22 | 1.69 | 0.52 | |
| N:P | 0.88*** | 1.67 | 1.23** | 1.77 | 0.90 | 1.64 | 1.54 | 2.00 | 3.66 | 2.21 | 0.83 | 1.56 | 1.04*** | 1.83 | 0.37*** | 1.48 | 0.68 | 1.63 | -1.46 | 1.02 | |
| SLA | STD | -1.21*** | 2.22 | -1.84 | 3.57 | 0.90 | -1.94 | -0.70 | 1.26 | -0.28 | 0.36 | 0.34 | -0.89 | -1.08*** | 1.96 | 1.21 | -2.57 | -1.13** | 2.03 | -3.66 | 6.85 |
| LNC | 0.82*** | -0.18 | 0.91 | -0.26 | 0.39 | 0.64 | 0.55 | 0.33 | 0.55 | 0.24 | -0.34 | 1.99 | 0.59*** | 0.30 | -0.70 | 2.91 | 0.69* | 0.11 | 0.89* | -0.26 | |
| LPC | 1.25*** | -2.42 | 1.51 | -2.81 | 0.96 | -1.87 | 1.12 | -2.18 | 2.79 | -5.58 | -0.81 | 1.53 | 1.31*** | -2.62 | -1.03 | 2.19 | 0.99* | -1.87 | 1.95* | -3.74 | |
| N:P | -0.85*** | 3.09 | -1.00 | 3.34 | -0.94 | 3.25 | 0.86 | -0.64 | -3.04 | 7.44 | 0.64 | 0.13 | -0.83*** | 3.14 | -0.40 | 2.16 | -0.63 | 2.68 | -1.67 | 4.68 | |
| STD | LA | -1.62*** | -1.11 | 0.75 | -0.84 | 1.20 | -0.83 | 1.80 | 0.36 | 2.33 | -0.25 | -1.46 | -1.32 | -1.31** | -0.96 | 0.44 | -0.90 | -0.63 | -0.67 | 0.56 | -0.37 |
| LNC | -0.57** | 1.35 | 0.49 | 1.62 | -0.44 | 1.36 | -0.78 | 1.31 | -0.58 | 1.21 | 0.64 | 1.46 | -0.54* | 1.36 | -0.57 | 1.42 | -0.37 | 1.43 | 0.24 | 1.55 | |
| LPC | -0.92* | -0.12 | 0.85 | 0.33 | 1.07 | 0.22 | -1.60 | -0.17 | -12.72 | -2.28 | 1.85 | 0.31 | -1.16** | -0.25 | -0.85 | 0 | -0.56 | -0.01 | 0.53 | 0.23 | |
| N:P | 0.62 | 1.54 | 0.55 | 1.41 | -1.05 | 1.21 | 1.23 | 1.60 | 12.63 | 3.58 | -1.54 | 1.08 | 0.75* | 1.65 | -0.33 | 1.31 | 0.36 | 1.50 | -0.46 | 1.27 | |
| LA | LNC | 0.38*** | 1.76 | 0.67 | 2.19 | 0.36 | 1.78 | 0.43 | 1.60 | -0.41* | 1.03 | 0.27 | 1.58 | 0.44*** | 1.78 | 1.29 | 2.81 | 0.74* | 1.88 | 0.43 | 1.71 |
| LPC | 0.57*** | 0.51 | 1.11 | 1.26 | 0.89 | 0.96 | -0.89 | 0.14 | 2.09 | 1.65 | 0.65* | 0.55 | 0.87*** | 0.59 | 1.91 | 2.04 | 1.05 | 0.65 | 0.94* | 0.57 | |
| N:P | -0.39* | 1.10 | 0.73 | 2.02 | 0.87 | 2.24 | 0.68 | 1.36 | -2.28 | -0.44 | -0.51 | 0.91 | -0.61*** | 1.06 | -0.74 | 0.64 | -0.67 | 1.08 | -0.81* | 0.97 | |
| LNC | LPC | 1.56*** | -2.21 | 1.66** | -2.38 | 2.46 | -3.44 | 2.05 | -2.86 | -5.05 | 6.87 | 2.40** | -3.24 | 2.24*** | -3.31 | 1.48*** | -2.11 | 1.42*** | -2.03 | 2.19* | -3.17 |
表3 内蒙古锦鸡儿属性状间的关系
Table 3 Correlations of traits for Caragana species in Nei Mongol, China
| x | y | 锦鸡儿属 C. species | 矮锦鸡儿 C. pygmaea | 矮脚锦鸡儿 C. brachypoda | 鬼箭锦鸡儿 C. jubata | 荒漠锦鸡儿 C. roborovskyi | 毛刺锦鸡儿 C. tibetica | 柠条锦鸡儿 C. korshinskii | 狭叶锦鸡儿 C. stenophylla | 小叶锦鸡儿 C. microphylla | 中间锦鸡儿 C. intermedia | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | 斜率 Slope | 截距 Intercept | ||
| h | LDMC | -0.31*** | 0.21 | 0.61 | -1.40 | 0.50* | -1.03 | -0.39 | 0.29 | -0.15** | -0.04 | -0.39 | 0.16 | -0.56*** | 0.80 | 0.86 | -1.66 | -0.45*** | 0.52 | -0.26 | 0.19 |
| SLA | 0.33*** | 1.45 | 0.75 | 0.70 | -0.47 | 2.69 | 0.70 | 0.81 | -0.18* | 2.27 | -0.51 | 2.43 | 0.69*** | 0.62 | -0.79 | 3.24 | 0.54*** | 1.00 | 0.23** | 1.56 | |
| STD | -0.43*** | 0.52 | -1.40 | 2.33 | 0.43 | -0.76 | -0.49 | 0.69 | 0.21 | -0.55 | -0.23 | -0.02 | -0.75** | 1.29 | 0.95 | -1.74 | -0.65*** | 0.97 | -0.83 | 1.16 | |
| LA | 0.64*** | -1.83 | 1.02 | -2.71 | 0.51 | -1.75 | -0.89 | 1.60 | 0.24* | -1.10 | 0.64* | -1.53 | 0.98*** | -2.64 | 0.42 | -1.66 | 0.38** | -1.21 | 0.47 | -1.36 | |
| LNC | 0.25*** | 1.06 | 0.68 | 0.37 | 0.19 | 1.15 | 0.38 | 0.77 | -0.10** | 1.49 | 0.17 | 1.16 | 0.42* | 0.65 | -0.54 | 2.41 | 0.26*** | 1.04 | 0.20 | 1.12 | |
| LPC | 0.37*** | -0.55 | 1.14 | -1.76 | -0.46 | 0.72 | 0.79 | -1.27 | 0.51 | -0.65 | 0.41** | -0.44 | 0.88*** | -1.74 | -0.80 | 1.45 | 0.36*** | -0.55 | 0.44 | -0.71 | |
| N:P | -0.26 | 1.82 | -0.75 | 2.65 | 0.45 | 0.72 | -0.60 | 2.44 | -0.55 | 2.06 | -0.33** | 1.69 | -0.58*** | 2.64 | 0.31 | 0.87 | -0.23 | 1.84 | 0.38 | 0.74 | |
| LDMC | SLA | -1.07*** | 1.67 | -1.24 | 1.58 | -0.95*** | 1.71 | -1.79 | 1.33 | 1.21 | 2.32 | -1.28 | 1.67 | -1.23*** | 1.60 | -0.92 | 1.71 | -1.20*** | 1.62 | -0.87 | 1.72 |
| STD | 1.32*** | 0.20 | 2.23 | 0.64 | -0.85 | -0.41 | -1.26 | -0.90 | -0.75 | -0.41 | -0.49 | -0.31 | 1.33*** | 0.22 | 1.11 | 0.11 | 1.35*** | 0.20 | 3.19 | 0.55 | |
| LA | -2.07*** | -1.39 | 1.69 | -0.34 | -1.03 | -1.32 | -2.27 | -1.26 | -1.61* | -1.16 | -1.51 | -1.24 | -1.74*** | -1.26 | 0.49 | -0.85 | -0.83* | -0.78 | 1.79 | -0.06 | |
| LNC | -0.86*** | 1.21 | -1.13 | 1.18 | -0.37 | 1.31 | -0.98 | 1.05 | 0.66** | 1.51 | -0.43 | 1.24 | -0.74*** | 1.23 | -0.64*** | 1.35 | -0.75*** | 1.27 | -0.77 | 1.27 | |
| LPC | -1.32*** | -0.33 | -1.88** | -0.43 | -0.91* | -0.22 | -2.01 | -0.69 | -3.36 | -0.78 | -1.04 | -0.27 | -1.69*** | -0.55 | -0.94*** | -0.11 | -1.06** | -0.22 | 1.69 | 0.52 | |
| N:P | 0.88*** | 1.67 | 1.23** | 1.77 | 0.90 | 1.64 | 1.54 | 2.00 | 3.66 | 2.21 | 0.83 | 1.56 | 1.04*** | 1.83 | 0.37*** | 1.48 | 0.68 | 1.63 | -1.46 | 1.02 | |
| SLA | STD | -1.21*** | 2.22 | -1.84 | 3.57 | 0.90 | -1.94 | -0.70 | 1.26 | -0.28 | 0.36 | 0.34 | -0.89 | -1.08*** | 1.96 | 1.21 | -2.57 | -1.13** | 2.03 | -3.66 | 6.85 |
| LNC | 0.82*** | -0.18 | 0.91 | -0.26 | 0.39 | 0.64 | 0.55 | 0.33 | 0.55 | 0.24 | -0.34 | 1.99 | 0.59*** | 0.30 | -0.70 | 2.91 | 0.69* | 0.11 | 0.89* | -0.26 | |
| LPC | 1.25*** | -2.42 | 1.51 | -2.81 | 0.96 | -1.87 | 1.12 | -2.18 | 2.79 | -5.58 | -0.81 | 1.53 | 1.31*** | -2.62 | -1.03 | 2.19 | 0.99* | -1.87 | 1.95* | -3.74 | |
| N:P | -0.85*** | 3.09 | -1.00 | 3.34 | -0.94 | 3.25 | 0.86 | -0.64 | -3.04 | 7.44 | 0.64 | 0.13 | -0.83*** | 3.14 | -0.40 | 2.16 | -0.63 | 2.68 | -1.67 | 4.68 | |
| STD | LA | -1.62*** | -1.11 | 0.75 | -0.84 | 1.20 | -0.83 | 1.80 | 0.36 | 2.33 | -0.25 | -1.46 | -1.32 | -1.31** | -0.96 | 0.44 | -0.90 | -0.63 | -0.67 | 0.56 | -0.37 |
| LNC | -0.57** | 1.35 | 0.49 | 1.62 | -0.44 | 1.36 | -0.78 | 1.31 | -0.58 | 1.21 | 0.64 | 1.46 | -0.54* | 1.36 | -0.57 | 1.42 | -0.37 | 1.43 | 0.24 | 1.55 | |
| LPC | -0.92* | -0.12 | 0.85 | 0.33 | 1.07 | 0.22 | -1.60 | -0.17 | -12.72 | -2.28 | 1.85 | 0.31 | -1.16** | -0.25 | -0.85 | 0 | -0.56 | -0.01 | 0.53 | 0.23 | |
| N:P | 0.62 | 1.54 | 0.55 | 1.41 | -1.05 | 1.21 | 1.23 | 1.60 | 12.63 | 3.58 | -1.54 | 1.08 | 0.75* | 1.65 | -0.33 | 1.31 | 0.36 | 1.50 | -0.46 | 1.27 | |
| LA | LNC | 0.38*** | 1.76 | 0.67 | 2.19 | 0.36 | 1.78 | 0.43 | 1.60 | -0.41* | 1.03 | 0.27 | 1.58 | 0.44*** | 1.78 | 1.29 | 2.81 | 0.74* | 1.88 | 0.43 | 1.71 |
| LPC | 0.57*** | 0.51 | 1.11 | 1.26 | 0.89 | 0.96 | -0.89 | 0.14 | 2.09 | 1.65 | 0.65* | 0.55 | 0.87*** | 0.59 | 1.91 | 2.04 | 1.05 | 0.65 | 0.94* | 0.57 | |
| N:P | -0.39* | 1.10 | 0.73 | 2.02 | 0.87 | 2.24 | 0.68 | 1.36 | -2.28 | -0.44 | -0.51 | 0.91 | -0.61*** | 1.06 | -0.74 | 0.64 | -0.67 | 1.08 | -0.81* | 0.97 | |
| LNC | LPC | 1.56*** | -2.21 | 1.66** | -2.38 | 2.46 | -3.44 | 2.05 | -2.86 | -5.05 | 6.87 | 2.40** | -3.24 | 2.24*** | -3.31 | 1.48*** | -2.11 | 1.42*** | -2.03 | 2.19* | -3.17 |
| h | LDMC | SLA | STD | LA | LNC | LPC | N:P | |
|---|---|---|---|---|---|---|---|---|
| 种内变异 Intraspecies variation | 46.11% | 20.42% | 25.91% | 26.05% | 26.49% | 14.26% | 29.82% | 21.19% |
| 种间变异 Interspecies variation | 70.54% | 20.55% | 21.36% | 19.66% | 75.46% | 19.59% | 22.89% | 12.72% |
表4 内蒙古锦鸡儿属植物功能性状在种内和种间的变异系数
Table 4 Intra- and interspecies variation coefficient for Caragana species functional traits in Nei Mongol, China
| h | LDMC | SLA | STD | LA | LNC | LPC | N:P | |
|---|---|---|---|---|---|---|---|---|
| 种内变异 Intraspecies variation | 46.11% | 20.42% | 25.91% | 26.05% | 26.49% | 14.26% | 29.82% | 21.19% |
| 种间变异 Interspecies variation | 70.54% | 20.55% | 21.36% | 19.66% | 75.46% | 19.59% | 22.89% | 12.72% |
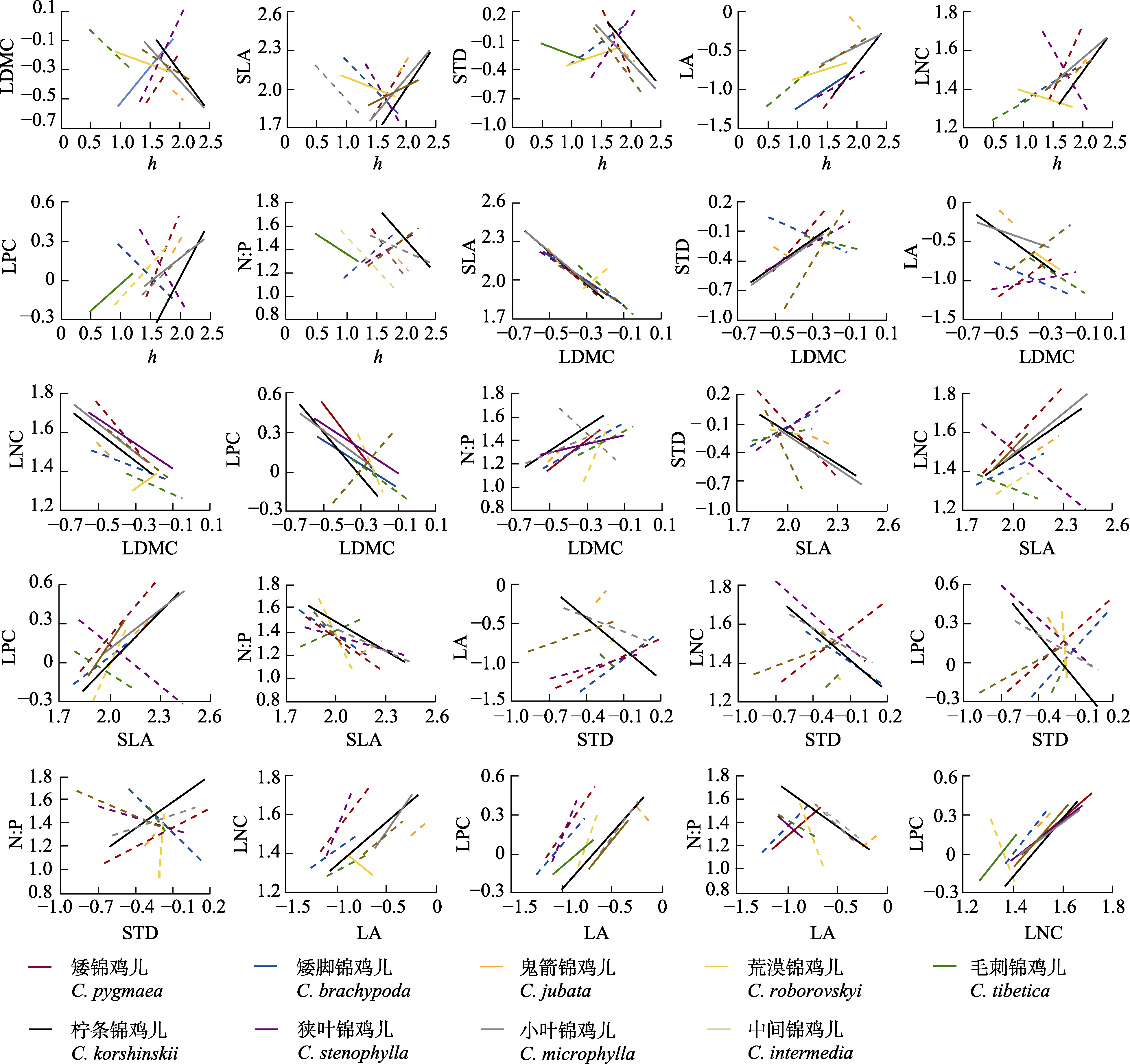
图4 内蒙古锦鸡儿属植物性状间的关系。实线表示p < 0.05, 虚线表示p ≥ 0.05。h, 株高; LA, 叶面积; LDMC, 叶干物质含量; LNC, 叶片氮含量; LPC, 叶片磷含量; N:P, 叶片氮磷比; SLA, 比叶面积; STD, 茎组织密度。
Fig. 4 Relationships between traits of Caragana species in Nei Mongol, China. Solid line represents p < 0.05, and dashed line represents p ≥ 0.05. h, height; LA, leaf area; LDMC, leaf dry matter content; LNC, leaf nitrogen content; LPC, leaf phosphorus content; N:P, leaf nitrogen content to phosphorus content ratio; SLA, specific leaf area; STD, stem tissue density.
| [1] |
Albert CH, Thuiller W, Yoccoz NG, Douzet R, Aubert S, Lavorel S (2010). A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Functional Ecology, 24, 1192-1201.
DOI URL |
| [2] |
Anderegg LDL, Berner LT, Badgley G, Sethi ML, Law BE, HilleRisLambers J (2018). Within-species patterns challenge our understanding of the leaf economics spectrum. Ecology Letters, 21, 734-744.
DOI PMID |
| [3] |
Blumenthal DM, Mueller KE, Kray JA, Ocheltree TW, Augustine DJ, Wilcox KR (2020). Traits link drought resistance with herbivore defence and plant economics in semi-arid grasslands: the central roles of phenology and leaf dry matter content. Journal of Ecology, 108, 2336-2351.
DOI URL |
| [4] |
Bowsher AW, Mason CM, Goolsby EW, Donovan LA (2016). Fine root tradeoffs between nitrogen concentration and xylem vessel traits preclude unified whole-plant resource strategies in Helianthus. Ecology and Evolution, 6, 1016-1031.
DOI PMID |
| [5] | Bruelheide H, Dengler J, Purschke O, Lenoir J, Jiménez-Alfaro B, Hennekens SM, Botta-Dukát Z, Chytrý M, Field R, Jansen F, Kattge J, Pillar VD, Schrodt F, Mahecha MD, Peet RK, et al. (2018). Global trait-environment relationships of plant communities. Nature Ecology & Evolution, 2, 1906-1917. |
| [6] |
Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, Zanne AE (2009). Towards a worldwide wood economics spectrum. Ecology Letters, 12, 351-366.
DOI PMID |
| [7] | de la Riva EG, Tosto A, Pérez-Ramos IM, Navarro-Fernández CM, Olmo M, Anten NPR, Maranon T, Villar R (2016). A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? Journal of Vegetation Science, 271, 87-199. |
| [8] |
Di Biase L, Fattorini S, Cutini M, Bricca A (2021). The role of inter-and intraspecific variations in grassland plant functional traits along an elevational gradient in a Mediterranean Mountain area. Plants, 10, 359. DOI: 10.3390/plants10020359.
DOI |
| [9] |
England JR, Attiwill PM (2006). Changes in leaf morphology and anatomy with tree age and height in the broadleaved evergreen species, Eucalyptus regnans F. Trees, 20, 79-90.
DOI URL |
| [10] |
Fick SE, Hijmans RJ (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37, 4302-4315.
DOI URL |
| [11] |
Freschet GT, Cornelissen JHC, Aerts R (2010). Evidence of the ‘plant economics spectrum’ in a subarctic flora. Journal of Ecology, 98, 362-373.
DOI URL |
| [12] |
Fyllas NM, Michelaki C, Galanidis A, Evangelou E, Zaragoza- Castells J, Dimitrakopoulos PG, Tsadilas C, Arianoutsou M, Lloyd J (2020). Functional trait variation among and within species and plant functional types in mountainous Mediterranean forests. Frontiers in Plant Science, 11, 212. DOI: 10.3389/fpls.2020.00212.
DOI |
| [13] |
Garnier E, Shipley B, Roumet C, Laurent G (2001). A standar- dized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology, 15, 688-695.
DOI URL |
| [14] |
Grime JP (1977). Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist, 111, 1169-1194.
DOI URL |
| [15] |
He NP, Li Y, Liu CC, Xu L, Li MX, Zhang JH, He JS, Tang ZY, Han XG, Ye Q, Xiao CW, Yu Q, Liu SR, Sun W, Niu SL, et al. (2020). Plant trait networks: improved resolution of the dimensionality of adaptation. Trends in Ecology & Evolution, 35, 908-918.
DOI URL |
| [16] |
Hinsinger P (2001). Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant and Soil, 237, 173-195.
DOI URL |
| [17] |
Jardine EC, Thomas GH, Forrestel EJ, Lehmann CER, Osborne CP (2020). The global distribution of grass functional traits within grassy biomes. Journal of Biogeography, 47, 553-565.
DOI URL |
| [18] |
Jin DM, Cao XC, Ma KP (2014). Leaf functional traits vary with the adult height of plant species in forest communities. Journal of Plant Ecology, 7, 68-76.
DOI URL |
| [19] | Kemppinen J, Niittynen P le Roux PC, Momberg M, Happonen K, Aalto J, Rautakoski H, Enquist BJ, Vandvik V, Halbritter AH, Maitner B, Luoto M (2021). Consistent trait-environment relationships within and across tundra plant communities. Nature Ecology & Evolution, 5, 458-467. |
| [20] |
Korall P, Pryer KM (2014). Global biogeography of scaly tree ferns (Cyatheaceae): evidence for Gondwanan vicariance and limited transoceanic dispersal. Journal of Biogeography, 41, 402-413.
PMID |
| [21] | Lambers H, Chapin III FS, Pons TL (1998). Plant Physiological Ecology. Spinger-Verlag, New York. |
| [22] |
Laughlin DC, Lusk CH, Bellingham PJ, Burslem DFRP, Simpson AH, Kramer-Walter KR (2017). Intraspecific trait variation can weaken interspecific trait correlations when assessing the whole-plant economic spectrum. Ecology and Evolution, 7, 8936-8949.
DOI PMID |
| [23] |
Liao JX, Chen J, Jiang MX, Huang HD (2012). Leaf traits and persistence of relict and endangered tree species in a rare plant community. Functional Plant Biology, 39, 512-518.
DOI PMID |
| [24] |
Liu MZ, Wang ZW,Li SS Lü X, Wang XB, Han XG (2017). Changes in specific leaf area of dominant plants in temperate grasslands along a 2500-km transect in Northern China. Scientific Reports, 7, 10780. DOI: 10.1038/s41598- 017-11133-z.
DOI |
| [25] | Liu XJ, Ma KP (2015). Plant functional traits—Concepts, applications and future directions. Scientia Sinica (Vitae), 45, 325-339. |
| [ 刘晓娟, 马克平 (2015). 植物功能性状研究进展. 中国科学: 生命科学, 45, 325-339.] | |
| [26] |
Lusk CH, Reich PB, Montgomery RA, Ackerly DD, Cavender-Bares J (2008). Why are evergreen leaves so contrary about shade? Trends in Ecology & Evolution, 23, 299-303.
DOI URL |
| [27] |
Meng H, Wei X, Franklin SB, Wu H, Jiang M (2017). Geographical variation and the role of climate in leaf traits of a relict tree species across its distribution in China. Plant Biology, 19, 552-561.
DOI PMID |
| [28] |
Messier J, McGill BJ, Enquist BJ, Lechowicz MJ (2017). Trait variation and integration across scales: Is the leaf economic spectrum present at local scales? Ecography, 40, 685-697.
DOI URL |
| [29] |
Niinemets Ü (2015). Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex. New Phytologist, 205, 79-96.
PMID |
| [30] |
Ordoñez JC, van Bodegom PM, Witte JPM, Wright IJ, Reich PB, Aerts R (2009). A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecology and Biogeography, 18, 137-149.
DOI URL |
| [31] |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE, Urcelay C, Veneklaas EJ, Reich PB, Poorter L, Wright IJ, et al. (2013). New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 61, 167-234.
DOI URL |
| [32] |
Pietsch KA, Ogle K, Cornelissen JHC, Cornwell WK, Bönisch G, Craine JM, Jackson BG, Kattge J, Peltzer DA, Penuelas J, Reich PB, Wardle DA, Weedon JT, Wright IJ, Zanne AE, Wirth C (2014). Global relationship of wood and leaf litter decomposability: the role of functional traits within and across plant organs. Global Ecology and Biogeography, 23, 1046-1057.
DOI URL |
| [33] |
Preston KA, Cornwell WK, DeNoyer JL (2006). Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms. New Phytologist, 170, 807-818.
PMID |
| [34] |
Reich PB, Ellsworth DS, Walters MB, Vose JM, Gresham C, Volin JC, Bowman WD (1999). Generality of leaf trait relationships: a test across six biomes. Ecology, 80, 1955-1969.
DOI URL |
| [35] |
Reich PB, Wright IJ, Lusk CH (2007). Predicting leaf physiology from simple plant and climate attributes: a global GLOPNET analysis. Ecological Applications, 17, 1982-1988.
DOI URL |
| [36] | Sack L, Cowan PD, Jaikumar N, Holbrook NM (2003). The “hydrology” of leaves: co-ordination of structure and function in temperate woody species. Plant, Cell & Environment, 26, 1343-1356. |
| [37] |
Stearns SC (1989). Trade-offs in life-history evolution. Functional Ecology, 3, 259-268.
DOI URL |
| [38] |
Sultan SE (2000). Phenotypic plasticity for plant development, function and life history. Trends in Plant Science, 5, 537-542.
DOI PMID |
| [39] |
Turtureanu PD, Barros C, Bec S, Hurdu BI, Saillard A, Šibík J, Balázs ZR, Novikov A, Renaud J, Podar D, Thuiller W, Pușcaș MH, Choler P (2020). Biogeography of intraspecific trait variability in matgrass (Nardus stricta): high phenotypic variation at the local scale exceeds large scale variability patterns. Perspectives in Plant Ecology, Evolution and Systematics, 46, 125555. DOI: 10.1016/j.ppees. 2020.125555
DOI |
| [40] |
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007). Let the concept of trait be functional! Oikos, 116, 882-892.
DOI URL |
| [41] |
Wei H, Wu B, Yang W, Luo T (2011). Low rainfall-induced shift in leaf trait relationship within species along a semi-arid sandy land transect in northern China. Plant Biology, 13, 85-92.
DOI PMID |
| [42] |
Westoby M (1998). A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil, 199, 213-227.
DOI URL |
| [43] |
Wilson PJ, Thompson K, Hodgson JG (1999). Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 143, 155-162.
DOI URL |
| [44] |
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, et al. (2004). The worldwide leaf economics spectrum. Nature, 428, 821-827.
DOI URL |
| [45] |
Xie LN, Ma CC, Guo HY, Li QF, Gao YB (2014). Distribution pattern of Caragana species under the influence of climate gradient in the Inner Mongolia region, China. Journal of Arid Land, 6, 311-323.
DOI URL |
| [46] | Yang SS, Wen ZM, Miao LP, Qi DH, Hua DW (2014). Responses of plant functional traits to micro-topographical changes in hilly and gully region of the Loess Plateau, China. Chinese Journal of Applied Ecology, 25, 3413-3419. |
| [ 杨士梭, 温仲明, 苗连朋, 戚德辉, 花东文 (2014). 黄土丘陵区植物功能性状对微地形变化的响应. 应用生态学报, 25, 3413-3419.] | |
| [47] |
Yang YZ, Wang H, Harrison SP, Prentice IC, Wright IJ, Peng CH, Lin GH (2019). Quantifying leaf-trait covariation and its controls across climates and biomes. New Phytologist, 221, 155-168.
DOI PMID |
| [48] | Zhou DW (1996). Study on distribution of the genus Caragana Fabr. Bulletin of Botanical Research, 16, 428-435. |
| [1] | 付粱晨, 丁宗巨, 唐茂, 曾辉, 朱彪. 北京东灵山白桦和蒙古栎的根际效应及其季节动态[J]. 植物生态学报, 2024, 48(4): 508-522. |
| [2] | 汤璐瑶, 方菁, 钱海蓉, 张博纳, 上官方京, 叶琳峰, 李姝雯, 童金莲, 谢江波. 落羽杉和池杉功能性状随高度的变异与协同[J]. 植物生态学报, 2023, 47(11): 1561-1575. |
| [3] | 张义, 程杰, 苏纪帅, 程积民. 长期封育演替下典型草原植物群落生产力与多样性关系[J]. 植物生态学报, 2022, 46(2): 176-187. |
| [4] | 祁鲁玉, 陈浩楠, 库丽洪·赛热别力, 籍天宇, 孟高德, 秦慧颖, 王宁, 宋逸欣, 刘春雨, 杜宁, 郭卫华. 基于植物功能性状的暖温带5种灌木幼苗生长策略[J]. 植物生态学报, 2022, 46(11): 1388-1399. |
| [5] | 严正兵, 刘树文, 吴锦. 高光谱遥感技术在植物功能性状监测中的应用与展望[J]. 植物生态学报, 2022, 46(10): 1151-1166. |
| [6] | 张景慧, 王铮, 黄永梅, 陈慧颖, 李智勇, 梁存柱. 草地利用方式对温性典型草原优势种植物功能性状的影响[J]. 植物生态学报, 2021, 45(8): 818-833. |
| [7] | 朱蔚娜, 张国龙, 张璞进, 张迁迁, 任瑾涛, 徐步云, 清华. 大针茅草原6种主要植物叶凋落物和根系分解特征与功能性状的关系[J]. 植物生态学报, 2021, 45(6): 606-616. |
| [8] | 王钊颖, 陈晓萍, 程英, 王满堂, 钟全林, 李曼, 程栋梁. 武夷山49种木本植物叶片与细根经济谱[J]. 植物生态学报, 2021, 45(3): 242-252. |
| [9] | 石娇星, 许洺山, 方晓晨, 郑丽婷, 张宇, 鲍迪峰, 杨安娜, 阎恩荣. 中国东部海岛黑松群落功能多样性的纬度变异及其影响因素[J]. 植物生态学报, 2021, 45(2): 163-173. |
| [10] | 潘权, 郑华, 王志恒, 文志, 杨延征. 植物功能性状对生态系统服务影响研究进展[J]. 植物生态学报, 2021, 45(10): 1140-1153. |
| [11] | 刘润红, 白金连, 包含, 农娟丽, 赵佳佳, 姜勇, 梁士楚, 李月娟. 桂林岩溶石山青冈群落主要木本植物功能性状变异与关联[J]. 植物生态学报, 2020, 44(8): 828-841. |
| [12] | 曹嘉瑜, 刘建峰, 袁泉, 徐德宇, 樊海东, 陈海燕, 谭斌, 刘立斌, 叶铎, 倪健. 森林与灌丛的灌木性状揭示不同的生活策略[J]. 植物生态学报, 2020, 44(7): 715-729. |
| [13] | 冯继广, 朱彪. 氮磷添加对树木生长和森林生产力影响的研究进展[J]. 植物生态学报, 2020, 44(6): 583-597. |
| [14] | 丁威,王玉冰,向官海,迟永刚,鲁顺保,郑淑霞. 小叶锦鸡儿灌丛化对典型草原群落结构与生态系统功能的影响[J]. 植物生态学报, 2020, 44(1): 33-43. |
| [15] | 王玉冰,孙毅寒,丁威,张恩涛,李文怀,迟永刚,郑淑霞. 长期氮添加对典型草原植物多样性与初级生产力的影响及途径[J]. 植物生态学报, 2020, 44(1): 22-32. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()