
植物生态学报 ›› 2018, Vol. 42 ›› Issue (2): 220-228.DOI: 10.17521/cjpe.2017.0258
所属专题: 植物功能性状
赵乐文1,陈梓熠1,邹滢1,付子钊1,吴桂林2,3,刘小容2,3,罗琦2,3,林忆雪4,2,李雄炬1,刘智通1,刘慧2,*( )
)
出版日期:2018-02-20
发布日期:2018-04-16
通讯作者:
刘慧
基金资助:
ZHAO Le-Wen1,CHEN Zi-Yi1,ZOU Ying1,FU Zi-Zhao1,WU Gui-Lin2,3,LIU Xiao-Rong2,3,LUO Qi2,3,LIN Yi-Xue4,2,LI Xiong-Ju1,LIU Zhi-Tong 1,LIU Hui2,*()
Online:2018-02-20
Published:2018-04-16
Contact:
Hui LIU
Supported by:摘要:
维管植物从蕨类植物、裸子植物到被子植物的演化过程中, 生理结构和功能都被认为更趋向于适应干旱的环境。但是关于3类植物是否提升了水分传导和水分利用效率的研究目前多基于叶脉和气孔的形态结构推断, 而对水分传导速率的直接测量则集中于被子植物内部。因此, 同时测定3类植物叶片和枝条尺度的水力性状, 可以研究维管植物水力性状的演化趋势和协同演化关系。该研究测定了中国科学院华南植物园内的蕨类植物(芒萁(Dicranopteris pedata)、华南毛蕨(Cyclosorus parasiticus)、乌毛蕨(Blechnum orientale)), 裸子植物(罗汉松(Podocarpus macrophyllus)、竹柏(Podocarpus nagi)、落羽杉(Taxodium distichum))和被子植物(海南木莲(Manglietia fordiana var. hainanensis)、东京油楠(Sindora tonkinensis)、羊蹄甲(Bauhinia purpurea))共9种植物的水力结构与功能的相关性状。发现9种植物从原始到更演化物种的枝条比导水率(KS)、叶片比导水率(KL)和叶片水力导度(Kleaf)依次升高; 叶片蒸腾速率(E)和潜在水分利用效率(WUEi)升高趋势不显著; 气孔面积指数(SPI)和木材密度(WD)在3个类群间无明显变化趋势。水力功能性状在枝条(KS)和叶片(KL、Kleaf和E)尺度显著相关, 但和两个结构性状并无显著相关性。对显著相关的性状进行系统发育独立差分析发现相关性依然存在, 表明枝条导水和叶片导水、叶片失水的协同性独立于系统发育而存在。该文以9种维管植物为例, 发现演化过程中水分传导相关的功能性状有显著提升, 且导水和失水的性状存在着协同演化关系。上述维管植物水力性状演化趋势的结论需要对更多物种、更精细的水力结构性状的进行测定验证。
赵乐文, 陈梓熠, 邹滢, 付子钊, 吴桂林, 刘小容, 罗琦, 林忆雪, 李雄炬, 刘智通, 刘慧. 九种维管植物水力性状的演化趋势. 植物生态学报, 2018, 42(2): 220-228. DOI: 10.17521/cjpe.2017.0258
ZHAOLe-Wen, CHEN Zi-Yi, ZOU Ying, FU Zi-Zhao, WU Gui-Lin, LIU Xiao-Rong, LUO Qi, LIN Yi-Xue, LI Xiong-Ju, LIU Zhi-Tong, LIU Hui. Changes in hydraulic traits of nine vascular plants from different evolutionary lineages. Chinese Journal of Plant Ecology, 2018, 42(2): 220-228. DOI: 10.17521/cjpe.2017.0258
| 物种 Species | 缩略词 Abbreviation | 科 Family | |
|---|---|---|---|
| 蕨类植物 Ferns | 芒萁 Dicranopteris pedata | Dp | 里白科 Gleicheniaceae |
| 华南毛蕨 Cyclosorus parasiticus | Cp | 金星蕨科 Thelypteridaceae | |
| 乌毛蕨 Blechnum orientale | Bo | 乌毛蕨科 Blechnaceae | |
| 裸子植物 Gymnosperms | 罗汉松 Podocarpus macrophyllus | Pm | 罗汉松科 Podocarpaceae |
| 竹柏 Podocarpus nagi | Pn | 罗汉松科 Podocarpaceae | |
| 落羽杉 Taxodium distichum | Td | 杉科 Taxodiaceae | |
| 被子植物 Angiosperms | 海南木莲 Manglietia fordiana var. hainanensis | Mh | 木兰科 Magnoliaceae |
| 东京油楠 Sindora tonkinensis | St | 豆科 Fabaceae | |
| 羊蹄甲 Bauhinia purpurea | Bp | 豆科 Fabaceae |
表1 本试验中不同演化类群9种维管植物概况
Table 1 Summary of the nine species from three evolutionary lineages in this study
| 物种 Species | 缩略词 Abbreviation | 科 Family | |
|---|---|---|---|
| 蕨类植物 Ferns | 芒萁 Dicranopteris pedata | Dp | 里白科 Gleicheniaceae |
| 华南毛蕨 Cyclosorus parasiticus | Cp | 金星蕨科 Thelypteridaceae | |
| 乌毛蕨 Blechnum orientale | Bo | 乌毛蕨科 Blechnaceae | |
| 裸子植物 Gymnosperms | 罗汉松 Podocarpus macrophyllus | Pm | 罗汉松科 Podocarpaceae |
| 竹柏 Podocarpus nagi | Pn | 罗汉松科 Podocarpaceae | |
| 落羽杉 Taxodium distichum | Td | 杉科 Taxodiaceae | |
| 被子植物 Angiosperms | 海南木莲 Manglietia fordiana var. hainanensis | Mh | 木兰科 Magnoliaceae |
| 东京油楠 Sindora tonkinensis | St | 豆科 Fabaceae | |
| 羊蹄甲 Bauhinia purpurea | Bp | 豆科 Fabaceae |

图1 不同演化类群9种植物的枝条比导水率(KS)、叶片比导水率(KL)和叶片水力导度(Kleaf)(平均值±标准误差, n = 3-5)。最右图柱上不同字母是HSD多重比较的结果, 9个物种的系统发育树在柱状图最下方, 物种缩写见表1。Fern, 蕨类植物; Gym, 裸子植物; Ang, 被子植物。
Fig. 1 Comparison of sapwood-specific hydraulic conductivity (KS), leaf-specific hydraulic conductivity (KL) and leaf hydraulic conductance (Kleaf) among the nine species from three evolutionary lineages (mean ± SE, n = 3-5). Letters on top of each bar in the right column are HSD multiple comparison results, the phylogenetic tree of the nine species is drawn at the bottom. See species abbrivations in Table 1. Fern, ferns; Gym, gynosperms; Ang, angiosperms.
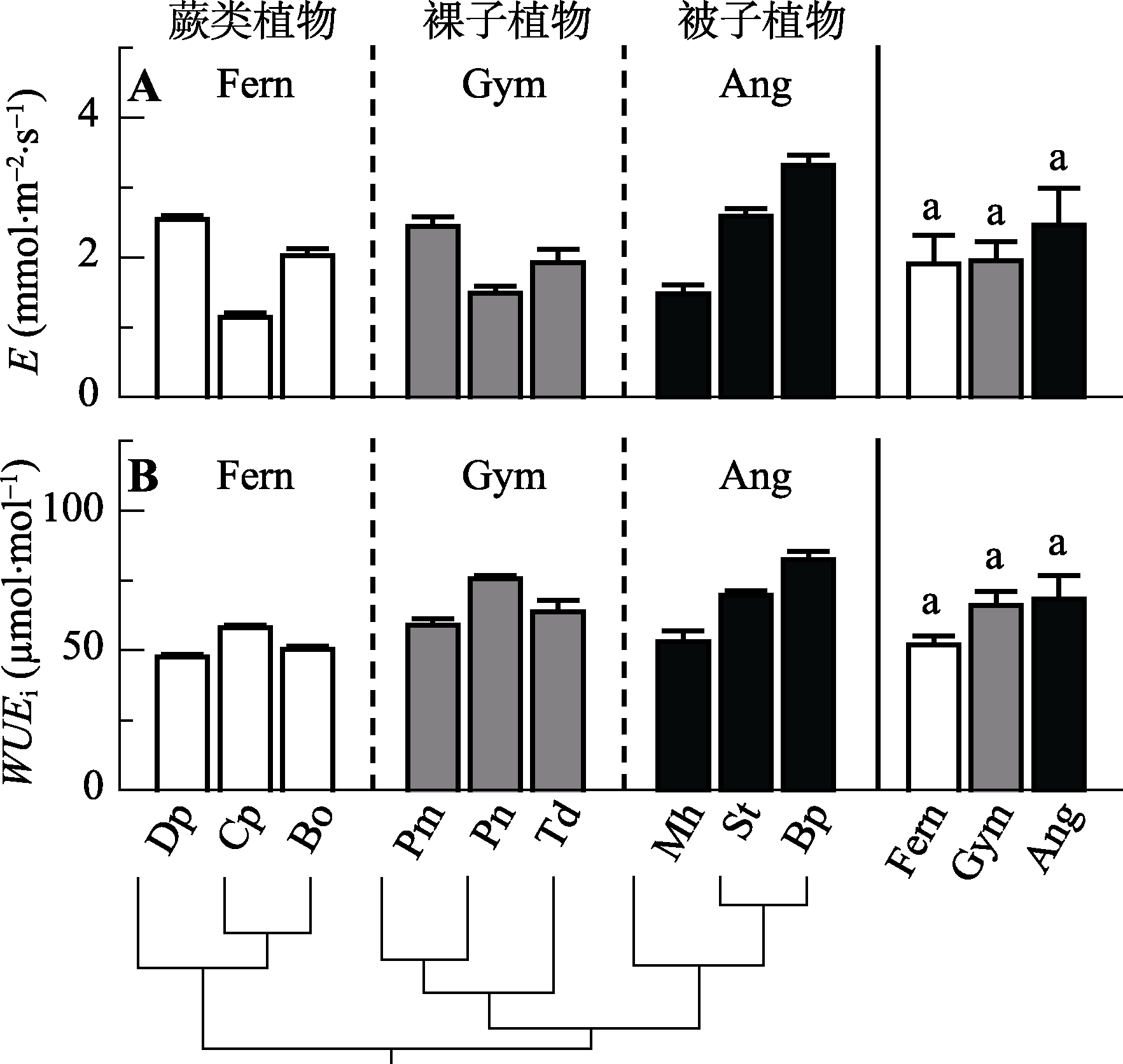
图2 不同演化类群9种植物的蒸腾速率(E)和潜在水分利用效率(WUEi)(平均值±标准误差, n = 3-5)。缩写注释同图1。
Fig. 2 Comparison of leaf transpiration rate (E) and intrinsic water use efficiency (WUEi) among the nine species from three evolutionary lineages (mean ± SE, n = 3-5). Notes are the same with Fig. 1.
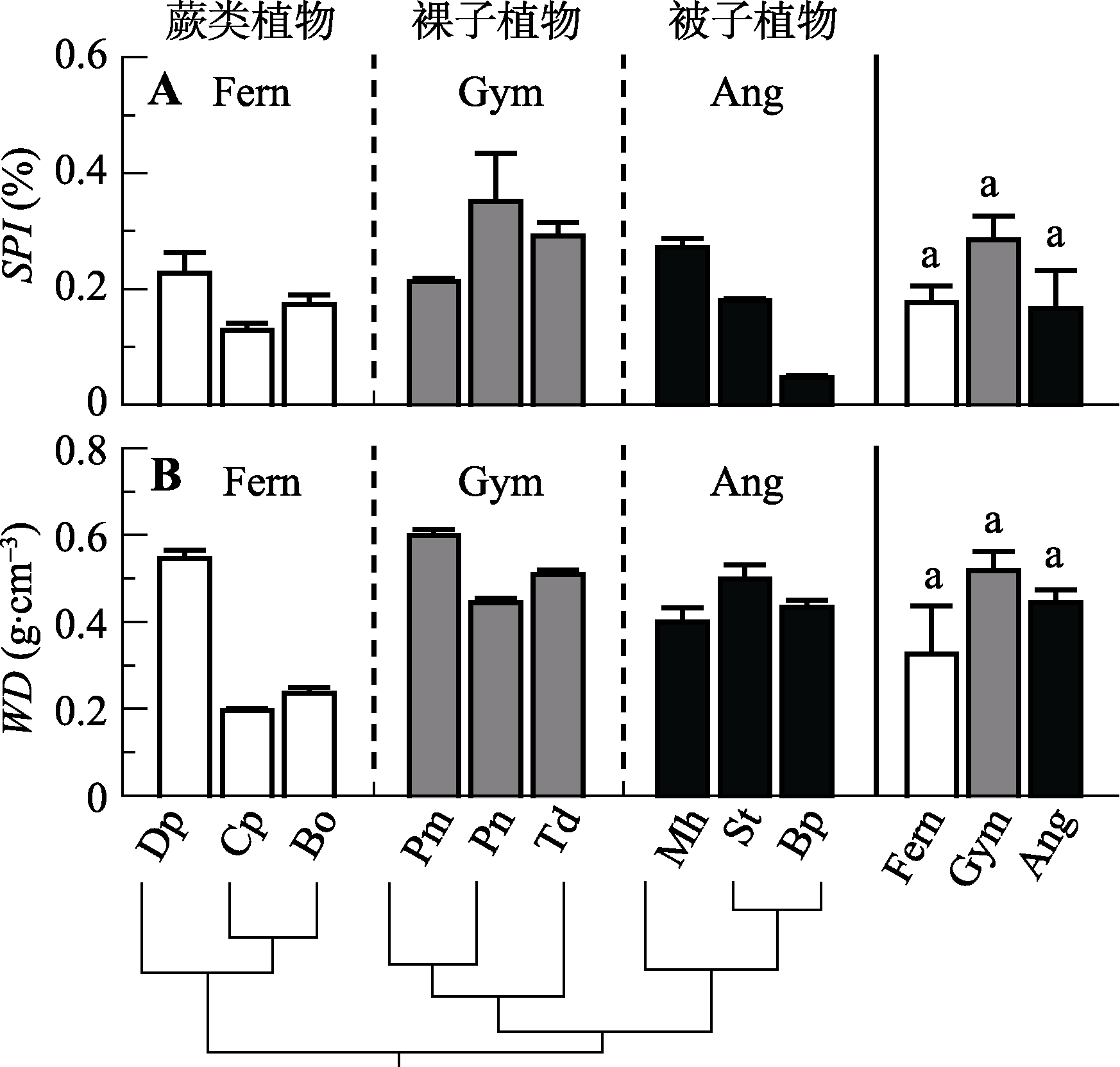
图3 不同演化类群9种植物的气孔面积指数(SPI)和木材密度(WD)(平均值±标准误差, n = 3-5)。缩写注释同图1。
Fig. 3 Comparison of stomatal area index (SPI) and wood density (WD) among the nine species from three evolutionary lineages (mean ± SE, n = 3-5). Notes are the same with Fig. 1.
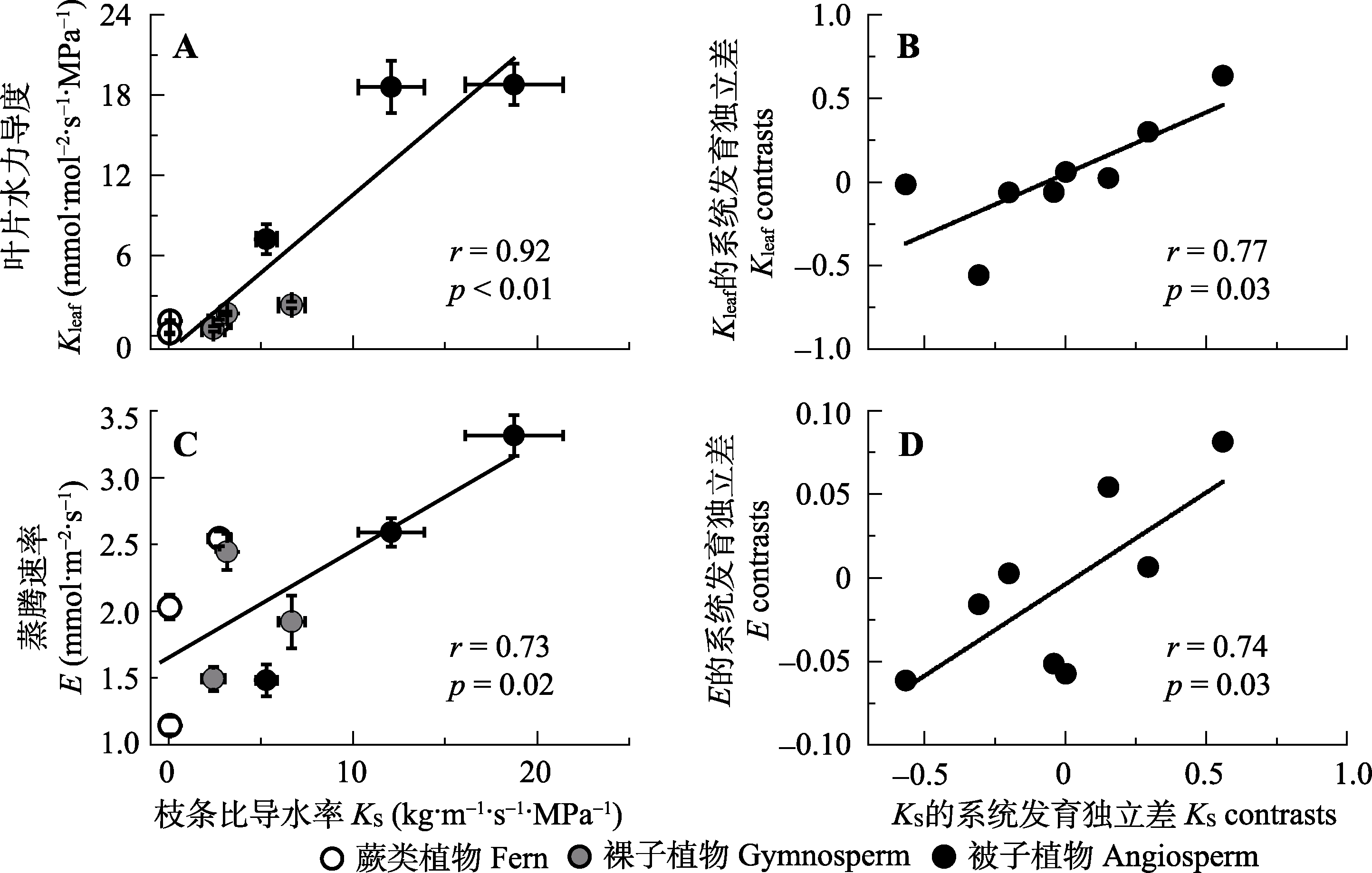
图4 不同演化类群9种植物的枝条比导水率(KS)与叶片水力导度(Kleaf)(A, B)和蒸腾速率(E)(C, D)的相关性。A、C是这两对性状的直接相关, B、D是这两对性状的系统发育独立差分析。A, C内数值为平均值±标准误差(n = 3-5), A, B, C, D均报告Pearson相关系数(r)和显著性(p)。
Fig. 4 Correlations between sapwood-specific hydraulic conductivity (KS) and leaf hydraulic conductance (Kleaf)(A, B), and leaf transpiration rate (E)(C, D) among the nine species from three evolutionary lineages. A, C, traditional cross-species correlations; B, D, correlations among phylogenetically independent contrasts. In A and C, data are mean ± SE (n = 3-5), Pearson correlation coefficients (r) and p values are reported.
| [1] |
Azani N, Babineau M, Bailey CD, Banks H, Barbosa AR, Pinto RB, Boatwright JS, Borges LM, Brown GK, Bruneau A, Candido E, Cardoso D, Chung K-F, Clark RP, Concei??o AdS, Crisp M, Cubas P, Delgado-Salinas A, Dexter KG, Doyle JJ, Duminil J, Egan AN, De La Estrella M, Falc?o MJ, Filatov DA, Fortuna-Perez AP, Fortunato RH, Gagnon E, Gasson P, Rando JG, Azevedo Tozzi AMGd, Gunn B, Harris D, Haston E, Hawkins JA, Herendeen PS, Hughes CE, Iganci JRV, Javadi F, Kanu SA, Kazempour- Osaloo S, Kite GC, Klitgaard BB, Kochanovski FJ, Koenen EJM, Kovar L, Lavin M, Roux Ml, Lewis GP, de Lima HC, López Roberts MC, Mackinder B, Maia VH, Malécot V, Mansano VF, Marazzi B, Mattapha S, Miller JT, Mitsuyuki C, Moura T, Murphy DJ, Nageswara-Rao M, Nevado B, Neves D, Ojeda DI, Pennington RT, Prado DE, Prenner G, de Queiroz LP, Ramos G, Ranzato Filardi FL, Ribeiro PG, Rico-Arce MdL, Sanderson MJ, Santos- Silva J, S?o-Mateus WMB, Silva MJS, Simon MF, Sinou C, Snak C, de Souza éR, Sprent J, Steele KP, Steier JE, Steeves R, Stirton CH, Tagane S, Torke BM, Toyama H, Cruz DTd, Vatanparast M, Wieringa JJ, Wink M, Wojciechowski MF, Yahara T, Yi T, Zimmerman E ( 2017). A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny, The Legume Phylogeny Working Group (LPWG). Taxon, 66, 44-77.
DOI URL |
| [2] |
Boyce CK, Brodribb TJ, Feild TS, Zwieniecki MA ( 2009). Angiosperm leaf vein evolution was physiologically and environmentally transformative. Proceedings of the Royal Society B, 276, 1771-1776.
DOI URL |
| [3] |
Brodribb TJ, Bienaimé D, Marmottant P ( 2016). Revealing catastrophic failure of leaf networks under stress. Proceedings of the National Academy of Sciences of the United States of America, 113, 4865-4869.
DOI URL PMID |
| [4] |
Brodribb TJ, Feild TS ( 2010). Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification. Ecology Letters, 13, 175-183.
DOI URL PMID |
| [5] |
Brodribb TJ, Feild TS, Jordan GJ ( 2007). Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiology, 144, 1890-1898.
DOI URL |
| [6] |
Brodribb TJ, Holbrook NM ( 2003 a). Changes in leaf hydraulic conductance during leaf shedding in seasonally dry tropical forest. New Phytologist, 158, 295-303.
DOI URL |
| [7] |
Brodribb TJ, Holbrook NM ( 2003 b). Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiology, 132, 2166-2173.
DOI URL |
| [8] |
Brodribb TJ, Holbrook NM, Zwieniecki MA, Palma B ( 2005). Leaf hydraulic capacity in ferns, conifers and angiosperms, impacts on photosynthetic maxima. New Phytologist, 165, 839-846.
DOI URL PMID |
| [9] |
Buckley LB, Kingsolver JG ( 2012). Functional and phylogenetic approaches to forecasting species responses to climate change. Annual Review of Ecology, Evolution, and Systematics, 43, 205-264.
DOI URL |
| [10] | Darwin C ( 1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray, London. |
| [11] |
Escalona JM, Flexas J, Medrano H ( 1999). Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Functional Plant Biology, 26, 421-433.
DOI URL |
| [12] |
Fan ZX, Zhang SB, Hao GY, Ferry Slik JW, Cao KF ( 2012). Hydraulic conductivity traits predict growth rates and adult stature of 40 Asian tropical tree species better than wood density. Journal of Ecology, 100, 732-741.
DOI URL |
| [13] |
Franks PJ, Beerling DJ ( 2009). Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proceedings of the National Academy of Sciences of the United States of America, 106, 10343-10347.
DOI URL |
| [14] |
Franks PJ, Drake PL, Beerling DJ ( 2009). Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density, an analysis using Eucalyptus globulus. Plant, Cell & Environment, 32, 1737-1748.
DOI URL PMID |
| [15] |
Freckleton RP, Harvey PH, Pagel M ( 2002). Phylogenetic analysis and comparative data, a test and review of evidence. The American Naturalist, 160, 712-726.
DOI URL PMID |
| [16] | Givnish TJ ( 2002). Adaptive significance of evergreen vs. deciduous leaves, solving the triple paradox. Silva Fennica, 36, 703-743. |
| [17] | Guyot G, Scoffoni C, Sack L ( 2012). Combined impacts of irradiance and dehydration on leaf hydraulic conductance, insights into vulnerability and stomatal control. Plant, Cell & Environment, 35, 857-871. |
| [18] |
Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloh KA ( 2001). Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia, 126, 457-461.
DOI URL |
| [19] |
Ishida A, Nakano T, Yazaki K, Matsuki S, Koike N, Lauenstein D, Shimizu M, Yamashita N ( 2008). Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms. Oecologia, 156, 193-202.
DOI URL PMID |
| [20] |
Liu H, Xu QY, He PC, Santiago LS, Yang KM, Ye Q ( 2015 a). Strong phylogenetic signals and phylogenetic niche conservatism in ecophysiological traits across divergent lineages of Magnoliaceae. Scientific Reports, 5, 12246. DOI: 10.1038/srep12246.
DOI URL PMID |
| [21] |
Liu YY, Song J, Wang M, Li N, Niu CY, Hao GY ( 2015 b). Coordination of xylem hydraulics and stomatal regulation in keeping the integrity of xylem water transport in shoots of two compound-leaved tree species. Tree Physiology, 35, 1333-1342.
DOI URL PMID |
| [22] |
Nicotra AB, Davidson A ( 2010). Adaptive phenotypic plasticity and plant water use. Functional Plant Biology, 37, 117-127.
DOI URL |
| [23] |
Nicotra AB, Leigh A, Boyce CK, Jones CS, Niklas KJ, Royer DL, Tsukaya H ( 2011). The evolution and functional significance of leaf shape in the angiosperms. Functional Plant Biology, 38, 535-552.
DOI URL |
| [24] |
Oren R, Sperry JS, Ewers BE, Pataki DE, Philips N, Megonigal JP ( 2001). Sensitivity of mean canopy stomatal conductance to vapor pressure deficit in a flooded Taxodium distichum L. forest: Hydraulic and non-hydraulic effects. Oecologia, 126, 21-29.
DOI URL PMID |
| [25] |
Paradis E, Claude J, Strimmer K ( 2004). APE, analyses of phylogenetics and evolution in R language. Bioinformatics, 20, 289-290.
DOI URL PMID |
| [26] | Quinn CJ, Price RA, Gadek PA ( 2002). Familial concepts and relationships in the conifer based on rbcL and matK sequence comparisons. Kew Bulletin, 57, 513-531. |
| [27] |
Sack L, Cowan PD, Jaikumar N, Holbrook NM ( 2003). The ‘hydrology’ of leaves, co-ordination of structure and function in temperature woody species. Plant, Cell & Environment, 26, 1343-1356.
DOI URL |
| [28] |
Santiago LS, Goldstein G, Meinzer FC, Fisher JB, Machado K, Woodruff D, Jones T ( 2004). Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia, 140, 543-550.
DOI URL PMID |
| [29] |
Scoffoni C, McKown AD, Rawls M, Sack L ( 2012). Dynamics of leaf hydraulic conductance with water status, quantification and analysis of species differences under steady state. Journal of Experimental Botany, 63, 643-658.
DOI URL PMID |
| [30] | Song J, Li RH, Zhu SD, Ye Q ( 2013). Leaf functional traits of ferns from different habitats in monsoon evergreen broad- leaved forest in Dinghushan Mountain . Journal of Tropical and Subtropical Botany, 21, 489-495. |
| 宋娟, 李荣华, 朱师丹 鼎湖山季风常绿阔叶林不同生境蕨类植物的叶片功能性状研究. 热带亚热带植物学报, 21, 489-495.] | |
| [31] |
Sperry JS, Donnelly JR, Tyree MT ( 1988). A method for measuring hydraulic conductivity and embolism in xylem. Plant, Cell & Environment, 11, 35-40.
DOI URL |
| [32] |
Testo W, Sundue M ( 2016). A 4000-species dataset provides new insight into the evolution of ferns. Molecular Phylogenetics and Evolution, 105, 200-211.
DOI URL PMID |
| [33] |
Trifiló P, Raimondo F, Savi T, Lo Gullo MA, Nardini A ( 2016). The contribution of vascular and extra-vascular water pathways to drought-induced decline of leaf hydraulic conductance. Journal of Experimental Botany, 67, erw268. DOI: 10.1093/jxb/erw268.
DOI URL PMID |
| [34] |
Tuzet A, Perrier A, Leuning R ( 2003). A coupled model of stomatal conductance, photosynthesis and transpiration. Plant, Cell & Environment, 26, 1097-1116.
DOI URL |
| [35] |
Webb CO, Ackerly DD, Kembel SW ( 2008). Phylocom, software for the analysis of phylogenetic community structure trait evolution. Bioinformatics, 24, 2098-2100.
DOI URL |
| [36] | Wu DL ( 2006). loral of Guangdong . Guangdong Science & Technology Press, Guangzhou. |
| [ 吴德邻 ( 2006). 广东植物志 .广东科技出版社, 广州.] | |
| [37] | Zhan XY, Yu GR, Sheng WP, Fang HJ ( 2012). Foliar water use efficiency and nitrogen use efficiency of dominant plant species in main forests along the North-South Transect of East China. Chinese Journal of Applied Ecology, 23, 587-594. |
| [ 展小云, 于贵瑞, 盛文萍, 方华军 ( 2012). 中国东部南北样带森林优势植物叶片的水分利用效率和氮素利用效率. 应用生态学报 , 23, 587-594.] | |
| [38] |
Zhang Y, Yang SJ, Sun M, Cao KF ( 2014). Stomatal traits are evolutionarily associated with vein density in basal angiosperms. Plant Science Journal, 32, 320-328.
DOI URL |
|
[ 张亚, 杨石建, 孙梅, 曹坤芳 ( 2014). 基部被子植物气孔性状与叶脉密度的关联演化.植物科学学报 , 32, 320-328.]
DOI URL |
| [1] | 刘瑶 钟全林 徐朝斌 程栋梁 郑跃芳 邹宇星 张雪 郑新杰 周云若. 不同大小刨花楠细根功能性状与根际微环境关系[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 徐子怡 金光泽. 阔叶红松林不同菌根类型幼苗细根功能性状的变异与权衡[J]. 植物生态学报, 2024, 48(5): 612-622. |
| [3] | 常晨晖 朱彪 朱江玲 吉成均 杨万勤. 森林粗木质残体分解研究进展[J]. 植物生态学报, 2024, 48(5): 541-560. |
| [4] | 付粱晨, 丁宗巨, 唐茂, 曾辉, 朱彪. 北京东灵山白桦和蒙古栎的根际效应及其季节动态[J]. 植物生态学报, 2024, 48(4): 508-522. |
| [5] | 范宏坤, 曾涛, 金光泽, 刘志理. 小兴安岭不同生长型阔叶植物叶性状变异及权衡[J]. 植物生态学报, 2024, 48(3): 364-376. |
| [6] | 刘聪聪, 何念鹏, 李颖, 张佳慧, 闫镤, 王若梦, 王瑞丽. 宏观生态学中的植物功能性状研究: 历史与发展趋势[J]. 植物生态学报, 2024, 48(1): 21-40. |
| [7] | 陈昭铨, 王明慧, 胡子涵, 郎学东, 何云琼, 刘万德. 云南普洱季风常绿阔叶林幼苗的群落构建机制[J]. 植物生态学报, 2024, 48(1): 68-79. |
| [8] | 袁雅妮, 周哲, 陈彬洲, 郭垚鑫, 岳明. 基于功能性状的锐齿槲栎林共存树种生态策略差异[J]. 植物生态学报, 2023, 47(9): 1270-1277. |
| [9] | 孙佳慧, 史海兰, 陈科宇, 纪宝明, 张静. 植物细根功能性状的权衡关系研究进展[J]. 植物生态学报, 2023, 47(8): 1055-1070. |
| [10] | 赵孟娟, 金光泽, 刘志理. 阔叶红松林3种典型蕨类叶功能性状的垂直变异[J]. 植物生态学报, 2023, 47(8): 1131-1143. |
| [11] | 冯珊珊, 黄春晖, 唐梦云, 蒋维昕, 白天道. 细叶云南松针叶形态和显微性状地理变异及其环境解释[J]. 植物生态学报, 2023, 47(8): 1116-1130. |
| [12] | 代景忠, 白玉婷, 卫智军, 张楚, 辛晓平, 闫玉春, 闫瑞瑞. 羊草功能性状对施肥的动态响应[J]. 植物生态学报, 2023, 47(7): 943-953. |
| [13] | 周莹莹, 林华. 不同水热梯度下冠层优势树种叶片热力性状及适应策略的变化趋势[J]. 植物生态学报, 2023, 47(5): 733-744. |
| [14] | 陈雪纯, 刘虹, 朱少琦, 孙铭遥, 宇振荣, 王庆刚. 漓江流域不同弃耕年限下4种常见草本植物功能性状种内变化及其影响因素[J]. 植物生态学报, 2023, 47(4): 559-570. |
| [15] | 王文伟, 韩伟鹏, 刘文文. 滨海湿地入侵植物互花米草叶片功能性状对潮位的短期响应[J]. 植物生态学报, 2023, 47(2): 216-226. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19