
植物生态学报 ›› 2016, Vol. 40 ›› Issue (6): 620-630.DOI: 10.17521/cjpe.2015.0449
所属专题: 生态化学计量
周正虎, 王传宽*( )
)
收稿日期:2015-12-07
接受日期:2016-03-26
出版日期:2016-06-30
发布日期:2016-06-15
通讯作者:
王传宽
基金资助:
Zheng-Hu ZHOU, Chuan-Kuan WANG*()
Received:2015-12-07
Accepted:2016-03-26
Online:2016-06-30
Published:2016-06-15
Contact:
Chuan-Kuan WANG
摘要:
微生物分解者的生存和生长策略、群落结构和功能会随着底物化学计量特征而改变, 从而强烈地影响底物的分解速度和元素的周转速率, 进而影响生态系统的功能过程。深入理解微生物生理代谢活动和群落结构与陆地生态系统过程之间的联系及其对全球变化的响应和反馈机理是生态学和全球变化生物学领域值得关注的重大科学命题。该文基于生态化学计量学理论和代谢理论, 首先介绍微生物在陆地生态系统碳氮磷循环中的作用; 然后综述微生物对分解底物化学计量变异性的响应和调节的4种主要机制: (1)调整微生物自身化学计量特征; (2)调整微生物群落结构; (3)产生特定的胞外酶以获取受限制的资源; (4)改变碳氮磷元素利用效率。最后, 通过分析当前研究不足, 提出该领域亟需关注的科学问题有: (1)综合阐明微生物对底物化学计量变化响应的各种机制及其相对重要性; (2)探索全球变化对微生物驱动的碳氮磷循环的影响; (3)探索微生物对底物化学计量变化适应对策的时空变化。
周正虎, 王传宽. 微生物对分解底物碳氮磷化学计量的响应和调节机制. 植物生态学报, 2016, 40(6): 620-630. DOI: 10.17521/cjpe.2015.0449
Zheng-Hu ZHOU, Chuan-Kuan WANG. Responses and regulation mechanisms of microbial decomposers to substrate carbon, nitrogen, and phosphorus stoichiometry. Chinese Journal of Plant Ecology, 2016, 40(6): 620-630. DOI: 10.17521/cjpe.2015.0449
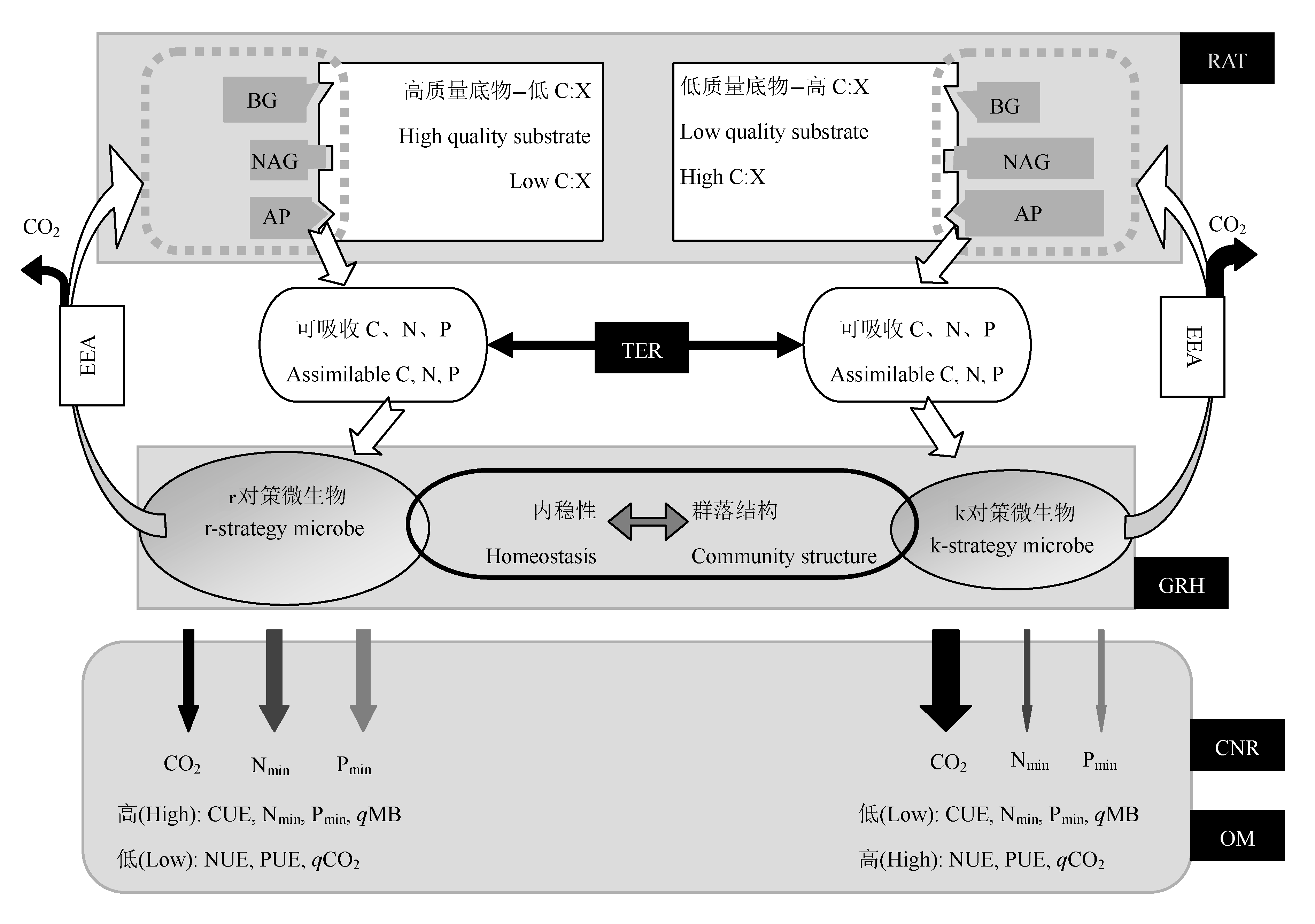
图1 微生物分解者对底物化学计量的响应和调节机制。C:X, 底物碳和养分(氮和磷)比; BG, β-1,4-葡萄糖苷酶; NAG, β-1,4-N-乙酰葡糖氨糖苷酶; AP, 磷酸酶; EEA, 胞外酶活性; RAT, 资源分配理论; TER, 元素比率阈值; GRH, 生长速率理论; CNR, 消费者驱动的养分循环; OM, 溢出代谢; CUE, 碳利用效率; NUE, 氮利用效率; PUE, 磷利用效率; qMB, 微生物生物量碳在底物总有机碳中的比例; qCO2, 单位微生物生物量碳呼吸速率; Nmin, 氮矿化; Pmin, 磷矿化。虚线方框中多边形大小表示胞外酶活大小; 椭圆大小表示微生物生物量大小; 箭头的宽窄表示碳氮磷通量大小。
Fig. 1 Mechanisms of microbial response and regulation on substrate stoichiometry. C:X, substrate carbon to nutrient (nitrogen and phosphor) ratios; BG, β-1,4-glucosidase; NAG, β-1,4-N-acetylglucosaminidase; AP, phosphatase, EEA, extracellular enzymatic activity; RAT, resource allocation theory; TER, threshold elemental ratio; GRH, growth rate hypothesis; CNR, consumer-driven nutrient recycling; OM, overflow metabolism; CUE, carbon use efficiency; NUE, nitrogen use efficiency; PUE, phosphor use efficiency; qMB, fraction of total substrate carbon in microbe; qCO2, microbial respiration rate per unit biomass carbon; Nmin, nitrogen mineralization; Pmin, phosphor mineralization. Size of the polygons in the dotted box indicates the strength of enzymatic activity; size of the ellipses indicates the amount of microbial biomass; and the width of the arrows indicates the magnitude of the carbon, nitrogen, and phosphor fluxes.
| [1] | Adamek M, Corre MD, Hölscher D (2009). Early effect of elevated nitrogen input on above-ground net primary production of a lower montane rain forest, Panama.Journal of Tropical Ecology, 25, 637-647. |
| [2] | Allison SD, Vitousek PM (2005). Responses of extracellular enzymes to simple and complex nutrient inputs.Soil Biology & Biochemistry, 37, 937-944. |
| [3] | Allison SD, Wallenstein MD, Bradford MA (2010). Soil-carbon response to warming dependent on microbial physiology.Nature Geoscience, 3, 336-340. |
| [4] | Barberán A, McGuire KL, Wolf JA, Jones FA, Wright SJ, Turner BL, Fierer N (2015). Relating belowground microbial composition to the taxonomic phylogenetic and functional trait distributions of trees in a tropical forest.Ecology Letters, 18, 1397-1405. |
| [5] | Bardgett RD, Freeman C, Ostle NJ (2008). Microbial contributions to climate change through carbon cycle feedbacks.The ISME Journal, 2, 805-814. |
| [6] | Blagodatskaya E, Kuzyakov Y (2013). Active microorganisms in soil: Critical review of estimation criteria and approaches.Soil Biology & Biochemistry, 67,192-211. |
| [7] | Bond-Lamberty B, Wang CK, Gower ST (2004). A global relationship between the heterotrophic and autotrophic components of soil respiration.Global Change Biology, 10, 1756-1766. |
| [8] | Borer ET, Bracken MES, Seabloom EW, Smith JE, Cebrian J, Cleland EE, Elser JJ, Fagan WF, Gruner DS, Harpole WS, Hillebrand H, Kerkhoff AJ, Ngai JT (2013). Global biogeography of autotroph chemistry: Is insolation a driving force?Oikos, 122, 1121-1130. |
| [9] | Brown JH, Gillooly JF, Allen AP, Savage VM, West GB (2004). Toward a metabolic theory of ecology.Ecology, 85, 1771-1789. |
| [10] | Buchkowski RW, Schmitz OJ, Bradford MA (2015). Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling.Ecology, 964, 1139-1149. |
| [11] | Cebrian J, Lartigue J (2004). Patterns of herbivory and decomposition in aquatic and terrestrial ecosystems.Ecological Monographs, 74, 237-259. |
| [12] | Chapin CT, Bridgham SD, Pastor J, Updegraff K (2003). Nitrogen phosphorus and carbon mineralization in response to nutrient and lime additions in peatlands.Soil Science, 168, 409-420. |
| [13] | Cleveland CC, Liptzin D (2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass?Biogeochemistry, 85, 235-252. |
| [14] | del Giorgio PA, Cole JJ (1998). Bacterial growth efficiency in natural aquatic systems.Annual Review of Ecology & Systematics, 29, 503-541. |
| [15] | Deng Q, Cheng XL, Yang YH, Zhang QF, Luo YQ (2014). Carbon-nitrogen interactions during afforestation in central China.Soil Biology & Biochemistry, 69, 119-122. |
| [16] | Doi H, Cherif M, Iwabuchi T, Katano I, Stegen JC, Striebel M (2010). Integrating elements and energy through the metabolic dependencies of gross growth efficiency and the threshold elemental ratio. Oikos, 119, 752-765. |
| [17] | Elser JJ (2006). Biological stoichiometry: A chemical bridge between ecosystem ecology and evolutionary biology.The American Naturalist, 168, S25-S35. |
| [18] | Elser JJ, Acharya K, Kyle M, Cotner J, Makino W, Markow T, Watts T, Hobbie S, Fagan W, Schade J, Hood J, Sterner RW (2003). Growth rate-stoichiometry couplings in diverse biota.Ecology Letters, 6, 936-943. |
| [19] | Fanin N, Fromin N, Buatois B, Hättenschwiler S (2013). An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system.Ecology Letters, 16, 764-772. |
| [20] | Fierer N, Bradford MA, Jackson RB (2007). Toward an ecological classification of soil bacteria.Ecology, 88, 1354-1364. |
| [21] | Fisk M, Santangelo S, Minick K (2015). Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests.Soil Biology & Biochemistry, 81, 212-218. |
| [22] | Frost PC, Benstead JP, Cross WF, Hillebrand H, Larson JH, Xenopoulos MA, Yoshida T (2006). Threshold elemental ratios of carbon and phosphorus in aquatic consumers.Ecology Letters, 9, 774-779. |
| [23] | Goberna M, Sánchez J, Pascual JA, García C (2006). Surface and subsurface organic carbon, microbial biomass and activity in a forest soil sequence.Soil Biology & Biochemistry, 38, 2233-2243. |
| [24] | Han WX, Tang LY, Chen YH, Fang JY (2013). Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants.PLoS ONE, 8, e83366 |
| [25] | Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD, Anderson NH, Cline SP, Aumen NG, Sedell JR, Lienkaemper GW, Cromack KJ, Cummins KW (1986). Ecology of coarse woody debris in temperate ecosystems.Advances in Ecological Research, 15, 133-302. |
| [26] | Hartman WH, Richardson CJ (2013). Differential nutrient limitation of soil microbial biomass and metabolic quotients (qCO2): Is there a biological stoichiometry of soil microbes.PloS ONE, 8, e57127. |
| [27] | Hessen DO, Elser JJ, Sterner RW, Urabe J (2013). Ecological stoichiometry: An elementary approach using basic principles.Limnology & Oceanography, 58, 2219-2236. |
| [28] | Högberg P, Nordgren A, Buchmann N, Taylor AF, Ekblad A, Högberg MN, Read DJ (2001). Large-scale forest girdling shows that current photosynthesis drives soil respiration.Nature, 411, 789-792. |
| [29] | Ihsam H, Haselwandter K (1989). Metabolic quotient of the soil microflora in relation to plant succession.Oecologia, 79, 174-178. |
| [30] | Ilg K, Wellbrock N, Lux W (2009). Phosphorus supply and cycling at long-term forest monitoring sites in Germany.European Journal of Forest Research, 128, 483-492. |
| [31] | Jansson JK, Prosser JI (2013). Microbiology: The life beneath our feet.Nature, 494, 40-41. |
| [32] | Jetten MS (2008). The microbial nitrogen cycle.Environmental Microbiology, 10, 2903-2909. |
| [33] | Kaiser C, Franklin O, Dieckmann U, Richter A (2014). Microbial community dynamics alleviate stoichiometric constraints during litter decay.Ecology letters, 17, 680-690. |
| [34] | Lee SH, Jang I, Chae N, Choi T, Kang H (2013). Organic layer serves as a hotspot of microbial activity and abundance in Arctic tundra soils.Microbial ecology, 65, 405-414. |
| [35] | Leff JW, Jones SE, Prober SM, Barberán A, Borer ET, Firn JL, Fierer N (2015). Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe.Proceedings of the National Academy of Sciences of the United States of America, 112, 10967-10972. |
| [36] | Li P, Han WX, Yang YH, Fang JY (2014). Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems. Global Ecology and Biogeography, 23, 979-987. |
| [37] | Li R, Chang RY (2015). Effects of external nitrogen additions on soil organic carbon dynamics and the mechanism.Chinese Journal of Plant Ecology, 39, 1012-1020. (in Chinese with English abstract)[李嵘, 常瑞英 (2015). 土壤有机碳对外源氮添加的响应及其机制. 植物生态学报, 39, 1012-1020.] |
| [38] | Li Y, Wu JS, Liu SL, Shen JL, Huang DY, Su YR, Wei WW, Syers JK (2012). Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Global Biogeochemical Cycles, 26, GB4002. doi: 10.1029/2012GB004399. |
| [39] | Lovelock CE, Feller IC, Ball MC, Ellis J, Sorrell B (2007). Testing the growth rate vs. geochemical hypothesis for latitudinal variation in plant nutrients.Ecology Letters, 10, 1154-1163. |
| [40] | Lu XQ, Toda H, Ding FJ, Fang SZ, Yang WX, Xu HG (2014). Effect of vegetation types on chemical and biological properties of soils of karst ecosystems.European Journal of Soil Biology, 61, 49-57. |
| [41] | Makino W, Cotner JB, Sterner RW, Elser JJ (2003). Are bacteria more like plants or animals? Growth rate and resource dependence of bacterial C: N: P stoichiometry.Functional Ecology, 17, 121-130. |
| [42] | Manzoni S, Taylor P, Richter A, Porporato A, Ågren GI (2012). Environmental and stoichiometric controls on microbial carbon-use efficiency in soils.New Phytologist, 196, 79-91. |
| [43] | Manzoni S, Trofymow JA, Jackson RB, Porporato A (2010). Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter.Ecological Monographs, 80, 89-106. |
| [44] | Marklein AR, Houlton BZ (2012). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems.New Phytologist, 193, 696-704. |
| [45] | Martin KJ, Picioreanu C, Nerenberg R (2015). Assessing microbial competition in a hydrogen-based membrane biofilm reactor (MBfR) using multidimensional modeling.Biotechnology & Bioengineering, 112, 1843-1853. |
| [46] | Martinelli LA, Almeida S, Brown IF, Moreira MZ, Victoria RL, Filoso S, Ferreira CAC, Thomas WW (2000). Variation in nutrient distribution and potential nutrient losses by selective logging in a humid tropical forest of Rondonia, Brazil.Biotropica, 32, 597-613. |
| [47] | McGroddy ME, Daufresne T, Hedin LO (2004). Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios.Ecology, 85, 2390-2401. |
| [48] | Moore J, Macalady JL, Schulz MS, White AF, Brantley SL (2010). Shifting microbial community structure across a marine terrace grassland chronosequence, Santa Cruz, California.Soil Biology & Biochemistry, 42, 21-31. |
| [49] | Moore JA, Jiang J, Patterson CM, Mayes MA, Wang G, Classen AT (2015). Interactions among roots mycorrhizas and free-living microbial communities differentially impact soil carbon processes.Journal of Ecology, 103, 1442-1453. |
| [50] | Moorhead DL, Sinsabaugh RL (2006). A theoretical model of litter decay and microbial interaction.Ecological Monographs, 76, 151-174. |
| [51] | Mooshammer M, Wanek W, Schnecker J, Wild B, Leitner S, Hofhansl F, Blöchl A, Hämmerle I, Frank AH, Keiblinger KM, Zechmeister-Boltenstern S, Richter A (2012). Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter.Ecology, 93, 770-782. |
| [52] | Mooshammer M, Wanek W, Zechmeister-Boltenstern S, Richter A (2014). Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources.Frontiers in Microbiology, 5, 22. |
| [53] | Odum EP (1969). The strategy of ecosystem development.Science, 164, 262-270 |
| [54] | Prosser JI, Bohannan BJ, Curtis TP, Ellis RJ, Firestone MK, Freckleton RP, Young JPW (2007). The role of ecological theory in microbial ecology.Nature Reviews Microbiology, 5, 384-392. |
| [55] | Raiesi F, Beheshti A (2015). Microbiological indicators of soil quality and degradation following conversion of native forests to continuous croplands.Ecological Indicators, 50, 173-185. |
| [56] | Redfield A (1958). The biological control of chemical factors in the environment.American Scientist, 46, 205-221. |
| [57] | Reed SC, Townsend AR, Davidson EA, Cleveland CC (2012). Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytologist, 196, 173-180. |
| [58] | Richardson AE, Simpson RJ (2011). Soil microorganisms mediating phosphorus availability update on microbial phosphorus.Plant Physiology, 156, 989-996. |
| [59] | Rick WY, Thomas SM (2001). Microbial nitrogen cycles: Physiology genomics and applications.Current Opinion in Microbiology, 4, 307-312. |
| [60] | Sardans J, Peñuelas J (2012). The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system.Plant Physiology, 160, 1741-1761. |
| [61] | Sardans J, Rivas-Ubach A, Peñuelas J (2012). The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives.Biogeochemistry, 111, 1-39. |
| [62] | Saviozzi A, Levi-Minzi R, Cardelli R, Riffaldi R (2001). A comparison of soil quality in adjacent cultivated, forest and native grassland soils.Plant & Soil, 233, 251-259. |
| [63] | Schimel JP, Weintraub MN (2003). The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model.Soil Biology & Biochemistry, 35, 549-563. |
| [64] | Schneider T, Keiblinger KM, Schmid E, Sterflinger-Gleixner K, Ellersdorfer G, Roschitzki B, Richter A, Eberl L, Zechmeister-Boltenstern S, Riedel K (2012). Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions.ISME Journal, 6, 1749-1762. |
| [65] | Seeling B, Zasoski RJ (1993). Microbial effects in maintaining organic and inorganic solution phosphorus concentrations in a grassland topsoil.Plant & Soil, 148, 277-284. |
| [66] | Sinsabaugh R, Follstad Shah JJ (2012). Ecoenzymatic sto- ichiometry and ecological theory.Annual Review of Ecology, Evolution, & Systematics, 43, 313-343. |
| [67] | Sinsabaugh RL, Follstad Shah JJ (2011). Ecoenzymatic stoichiometry of recalcitrant organic matter decomposition: The growth rate hypothesis in reverse.Biogeochemistry, 102, 31-43. |
| [68] | Sinsabaugh RL, Hill BH, Follstad Shah JJ (2009). Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment.Nature, 462, 795-798. |
| [69] | Sinsabaugh RL, Lauber CL, Weintraub MN, Ahmed B, Allison SD, Crenshaw C, Contosta AR, Cusack D, Frey S, Gallo ME (2008). Stoichiometry of soil enzyme activity at global scale. Ecology Letters, 11, 1252-1264. |
| [70] | Sinsabaugh RL, Manzoni S, Moorhead DL, Richter A (2013). Carbon use efficiency of microbial communities: Stoichio- metry, methodology and modelling.Ecology Letters, 16, 930-939. |
| [71] | Spohn M (2015). Microbial respiration per unit microbial biomass depends on litter layer carbon-to-nitrogen ratio. Biogeosciences, 12, 817-823. |
| [72] | Spohn M, Chodak M (2015). Microbial respiration per unit biomass increases with carbon-to-nutrient ratios in forest soils.Soil Biology & Biochemistry, 81, 128-133. |
| [73] | Steinauer K, Tilman D, Wragg PD, Cesarz S, Cowles JM, Pritsch K, Reich PB, Weisser WW, Eisenhauer N (2015). Plant diversity effects on soil microbial functions and enzymes are stronger than warming in a grassland experiment.Ecology, 96, 99-112. |
| [74] | Sterner RW, Elser JJ (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton. |
| [75] | Strickland MS, Osburn E, Lauber C, Fierer N, Bradford MA (2009). Litter quality is in the eye of the beholder: Initial decomposition rates as a function of inoculum characteristics.Functional Ecology, 23, 627-636. |
| [76] | Strickland MS, Rousk J (2010). Considering fungal: Bacterial dominance in soils-methods, controls, and ecosystem implications.Soil Biology & Biochemistry, 42, 1385-1395. |
| [77] | Susyan EA, Wirth S, Ananyeva ND, Stolnikova EV (2011). Forest succession on abandoned arable soils in European Russia-Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity.European Journal of Soil Biology, 47, 169-174. |
| [78] | Tian HQ, Chen GS, Zhang C, Melillo JM, Hall CAS (2010). Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data.Biogeochemistry, 98, 139-151. |
| [79] | van der Heijden MG, Bardgett RD, van Straalen NM (2008). The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems.Ecology letters, 11, 296-310. |
| [80] | Vergutz L, Manzoni S, Porporato A, Novais RF, Jackson RB (2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants.Ecological Monographs, 82, 205-220. |
| [81] | Vitousek PM, Howarth RW (1991). Nitrogen limitation on land and in the sea: How can it occur?Biogeochemistry, 13, 87-115. |
| [82] | Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010). Terrestrial phosphorus limitation: Mechanisms implications and nitrogen-phosphorus interactions.Ecological Applications, 20, 5-15. |
| [83] | Wang CK, Han Y, Chen JQ, Wang XC, Zhang QZ, Bond-Lamberty B (2013). Seasonality of soil CO2 efflux in a temperate forest: Biophysical effects of snowpack and spring freeze-thaw cycles.Agricultural & forest meteorology, 177, 83-92. |
| [84] | Wang CK, Yang JY (2007). Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests.Global Change Biology, 13, 123-131. |
| [85] | Wang CK, Yang JY, Zhang QZ (2006). Soil respiration in six temperate forests in China.Global Change Biology, 12, 2103-2114. |
| [86] | Wang CS, Wang SP (2015). A review of research on responses of leaf traits to climate change. ChineseJournal of Plant Ecology, 39, 206-216. (in Chinese with English abstract)[王常顺, 汪诗平 (2015). 植物叶片性状对气候变化的响应研究进展. 植物生态学报, 39, 206-216.] |
| [87] | Wang XC, Wang CK (2015). Fundamental concepts and field measurement methods of carbon cycling in forest ecosystems: a review.Acta Ecologica Sinica, 35, 4241-4256. (in Chinese with English abstract)[王兴昌, 王传宽 (2015). 森林生态系统碳循环的基本概念和野外测定方法评述. 生态学报, 35, 4241-4256.] |
| [88] | Wardle DA, Ghani A (1995). A critique of the microbial metabolic quotient (qCO2) as a bioindicator of disturbance and ecosystem development.Soil Biology & Biochemistry, 27, 1601-1610. |
| [89] | Waring BG, Weintraub SR, Sinsabaugh RL (2014). Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils.Biogeochemistry, 117, 101-113. |
| [90] | Weedon JT, Cornwell WK, Cornelissen JHC, Zanne AE, Wirth C, Coomes DA (2009). Global meta-analysis of wood decomposition rates: A role for trait variation among tree species?Ecology Letters, 12, 45-56. |
| [91] | Xu XF, Schimel JP, Thornton PE, Song X, Yuan FM, Goswami S (2014). Substrate and environmental controls on microbial assimilation of soil organic carbon: A framework for Earth system models.Ecology Letters, 17, 547-555. |
| [92] | Xu XF, Thornton PE, Post WM (2013). A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems.Global Ecology & Biogeography, 22, 737-749. |
| [93] | Yang X, Huang Z, Zhang K, Cornelissen JH (2015). C:N:P stoichiometry of Artemisia species and close relatives across northern China: Unravelling effects of climate, soil and taxonomy.Journal of Ecology, 103, 1020-1031. |
| [94] | Yang YH, Luo YQ (2011). Carbon:nitrogen stoichiometry in forest ecosystems during stand development.Global Ecology & Biogeography, 20, 354-361. |
| [95] | Yang YH, Luo YQ, Finzi AC (2011). Carbon and nitrogen dynamics during forest stand development: A global synthesis.New Phytologist, 190, 977-989. |
| [96] | Yuan ZY, Chen HY (2009). Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Global Ecology & Biogeography, 18, 11-18. |
| [97] | Yuan ZY, Chen HY, Reich PB (2011). Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus.Nature Communications, 2, 344. |
| [98] | Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W (2015). The application of ecological stoichiometry to plant-microbial- soil organic matter transformations.Ecological Monographs, 85, 133-155. |
| [99] | Zelezniak A, Andrejev S, Ponomarova O, Mende DR, Bork P, Patil KR (2015). Metabolic dependencies drive species co-occurrence in diverse microbial communities.Proceedings of the National Academy of Sciences of the United States of America, 112, 6449-6454. |
| [100] | Zhang QZ, Wang CK (2010). Carbon density and distribution of six Chinese temperate forests. Science China (Life Science), 53, 831-840. |
| [101] | Zhou ZH, Wang CK (2015). Reviews and syntheses: Soil resources and climate jointly drive variations in microbial1)biomass carbon and nitrogen in China’s forest ecosystems.Biogeosciences, 12, 6751-6760. |
| [102] | Zhou ZH, Wang CK, Zhang QZ (2015). The effect of land use change on soil carbon, nitrogen, and phosphorus contents and their stoichiometry in temperate sapling stands in northeastern China.Acta Ecologica Sinica, 35, 6694-6702. (in Chinese with English abstract)[周正虎, 王传宽, 张全智 (2015). 土地利用变化对东北温带幼龄林土壤碳氮磷含量及其化学计量特征的影响. 生态学报, 35, 6694-6702.] |
| [1] | 刘瑶 钟全林 徐朝斌 程栋梁 郑跃芳 邹宇星 张雪 郑新杰 周云若. 不同大小刨花楠细根功能性状与根际微环境关系[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 张文瑾 佘维维 秦树高 乔艳桂 张宇清. 氮和水分添加对黑沙蒿群落优势植物叶片氮磷化学计量特征的影响[J]. 植物生态学报, 2024, 48(5): 590-600. |
| [3] | 付粱晨, 丁宗巨, 唐茂, 曾辉, 朱彪. 北京东灵山白桦和蒙古栎的根际效应及其季节动态[J]. 植物生态学报, 2024, 48(4): 508-522. |
| [4] | 吴君梅, 曾泉鑫, 梅孔灿, 林惠瑛, 谢欢, 刘苑苑, 徐建国, 陈岳民. 土壤磷有效性调控亚热带森林土壤酶活性和酶化学计量对凋落叶输入的响应[J]. 植物生态学报, 2024, 48(2): 242-253. |
| [5] | 韩路, 冯宇, 李沅楷, 王雨晴, 王海珍. 地下水埋深对灰胡杨叶片与土壤养分生态化学计量特征及其内稳态的影响[J]. 植物生态学报, 2024, 48(1): 92-102. |
| [6] | 黄鹏, 林勇文, 张杰, 姚锦爱, 余德亿. 榕属植物特定挥发物决定榕管蓟马的寄主选择行为[J]. 植物生态学报, 2023, 47(7): 954-966. |
| [7] | 郭敏, 罗林, 梁进, 王彦杰, 赵春章. 冻融变化对西南亚高山森林优势种云杉和华西箭竹根区土壤理化性质与酶活性的影响[J]. 植物生态学报, 2023, 47(6): 882-894. |
| [8] | 罗娜娜, 盛茂银, 王霖娇, 石庆龙, 何宇. 长期植被恢复对中国西南喀斯特石漠化土壤活性有机碳组分含量和酶活性的影响[J]. 植物生态学报, 2023, 47(6): 867-881. |
| [9] | 张琦, 冯可, 常智慧, 何双辉, 徐维启. 灌丛化对林草交错带植物和土壤微生物的影响[J]. 植物生态学报, 2023, 47(6): 770-781. |
| [10] | 郑炀, 孙学广, 熊洋阳, 袁贵云, 丁贵杰. 叶际微生物对马尾松凋落针叶分解的影响[J]. 植物生态学报, 2023, 47(5): 687-698. |
| [11] | 赵小祥, 朱彬彬, 田秋香, 林巧玲, 陈龙, 刘峰. 叶片凋落物分解的主场优势研究进展[J]. 植物生态学报, 2023, 47(5): 597-607. |
| [12] | 李兆光, 文高, 和桂青, 徐天才, 和琼姬, 侯志江, 李燕, 薛润光. 滇西北藜麦氮磷钾生态化学计量特征的物候期动态[J]. 植物生态学报, 2023, 47(5): 724-732. |
| [13] | 林少颖, 曾瑜, 杨文文, 陈斌, 阮敏敏, 尹晓雷, 阳祥, 王维奇. 添加秸秆及其生物炭对茉莉植株与土壤碳氮磷生态化学计量特征的影响[J]. 植物生态学报, 2023, 47(4): 530-545. |
| [14] | 刘婧, 缑倩倩, 王国华, 赵峰侠. 晋西北丘陵风沙区柠条锦鸡儿叶片与土壤生态化学计量特征[J]. 植物生态学报, 2023, 47(4): 546-558. |
| [15] | 张尧, 陈岚, 王洁莹, 李益, 王俊, 郭垚鑫, 任成杰, 白红英, 孙昊田, 赵发珠. 太白山不同海拔森林根际土壤微生物碳利用效率差异性及其影响因素[J]. 植物生态学报, 2023, 47(2): 275-288. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19