
植物生态学报 ›› 2016, Vol. 40 ›› Issue (4): 341-353.DOI: 10.17521/cjpe.2015.0192
所属专题: 碳储量
范春楠1,2, 韩士杰2, 郭忠玲1,*( ), 郑金萍1, 程岩3
), 郑金萍1, 程岩3
收稿日期:2015-06-02
接受日期:2015-10-24
出版日期:2016-04-29
发布日期:2016-04-30
通讯作者:
郭忠玲
基金资助:
Chun-Nan FAN1,2, Shi-Jie HAN2, Zhong-Ling GUO1,*(), Jin-Ping ZHENG1, Yan CHENG3
Received:2015-06-02
Accepted:2015-10-24
Online:2016-04-29
Published:2016-04-30
Contact:
Zhong-Ling GUO
摘要:
通过对吉林省森林植被的普遍调查、典型调查以及植被样品含碳率测定, 结合吉林省2009年和2014年森林清查数据, 估算了区域森林植被的碳储量、碳密度及固碳速率。研究结果表明: 林下植被的生物量在不同林分和同类林分中存在较大的差异, 整体不足乔木层生物量的3%, 灌木植物的生物量略高于草本植物和幼树。不同林分类型的乔木含碳率介于45.80%-52.97%之间, 整体表现为针叶林高于阔叶林; 灌木和草本植物分别为39.79%-47.25%和40%左右。吉林省森林植被碳转换系数以0.47或0.48更为准确, 若以0.50或0.45作为植被的碳转换系数计算碳储量, 会造成±5.26%的偏差。吉林省森林植被不仅维持着较高的碳库水平, 而且极具碳汇能力; 2009年和2014年碳储量分别为471.29 Tg C和505.76 Tg C, 累计碳增量34.47 Tg C, 平均每年碳增量6.89 Tg C·a-1; 碳密度由64.58 t·hm-2增至66.68 t·hm-2, 平均增加2.10 t·hm-2, 固碳速率0.92 t·hm-2·a-1。森林植被碳储量的增长主体是蒙古栎(Quercus mongolica)林和阔叶混交林, 合计碳增量占总体的90.34%。受植被发育引起的生物量增长、林分龄组晋级以及森林经营所引起的面积变化影响, 各龄组植被碳增量为幼龄林>过熟林>近熟林>中龄林, 成熟林表现为负增长; 固碳速率为过熟林>幼龄林>近熟林>中龄林>成熟林。森林植被碳储量和碳密度的市/区分布整体表现为自东向西明显的降低变化; 碳增量以东北和中东部地区较高, 西部地区较低; 固碳速率整体以南部的通化地区和白山地区相对较高, 中部的吉林地区和东部的延边地区次之, 西部的白城地区、松原地区等地呈负增长。
范春楠, 韩士杰, 郭忠玲, 郑金萍, 程岩. 吉林省森林植被固碳现状与速率. 植物生态学报, 2016, 40(4): 341-353. DOI: 10.17521/cjpe.2015.0192
Chun-Nan FAN, Shi-Jie HAN, Zhong-Ling GUO, Jin-Ping ZHENG, Yan CHENG. Present status and rate of carbon sequestration of forest vegetation in Jilin Province, Northeast China. Chinese Journal of Plant Ecology, 2016, 40(4): 341-353. DOI: 10.17521/cjpe.2015.0192
| 林分类型 Forest type | 林下植被生物量 Biomass of understory vegetation (t·hm-2) | 植被含碳率 Carbon content of vegetation (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 幼树 Young tree | 灌木 Shrub | 草本 Herb | 合计 Total | 乔木 Tree | 灌木 Shrub | 草本 Herb | ||
| 白桦林 Betula platyphylla forest | 1.07 (56.57) | 0.28 (10.71) | 0.63 (34.92) | 1.98 (1.27) | 48.22 (0.70) | 43.58 (4.92) | 40.76 (5.86) | |
| 山杨林 Populus davidiana forest | 0.52 (17.38) | 0.47 (17.02) | 1.34 (35.07) | 2.33 (1.24) | 48.45 (0.55) | 47.25 (1.13) | 42.16 (2.88) | |
| 胡桃楸林 Juglans mandshurica forest | 0.66 (81.86) | 0.42 (7.14) | 0.22 (22.73) | 1.30 (1.31) | 45.80 (0.74) | 42.58 (7.96) | 39.24 (5.52) | |
| 椴树林 Tilia forest | 0.31 (74.19) | 0.53 (20.75) | 0.99 (7.07) | 1.83 (1.38) | 46.77 (0.31) | 45.76 (5.44) | 41.51 (6.44) | |
| 蒙古栎林 Quercus mongolica forest | 0.47 (111.79) | 2.36 (32.63) | 0.68 (22.06) | 3.51 (3.31) | 48.46 (0.41) | 43.69 (4.35) | 37.24 (5.34) | |
| 杂木林 Undesirable tree forest | 0.25 (18.56) | 1.61 (22.36) | 0.70 (35.71) | 2.56 (2.87) | 46.36 (0.43) | 39.79 (7.74) | 42.66 (4.52) | |
| 阔叶混交林 Broadleaf mixed forest | 0.72 (83.30) | 2.26 (22.57) | 0.86 (12.79) | 3.84 (2.59) | 47.60 (2.43) | 43.02 (8.48) | 37.80 (6.88) | |
| 针阔混交林 Needle broad-leaved mixed forest | 0.99 (74.17) | 2.20 (15.00) | 1.26 (17.46) | 4.45 (2.22) | 49.51 (1.85) | 45.44 (6.00) | 40.68 (7.30) | |
| 针叶混交林 Needle mixed forest | 0.96 (111.88) | 0.47 (17.02) | 0.80 (13.75) | 2.23 (1.29) | 51.08 (1.08) | 45.67 (3.32) | 42.62 (2.53) | |
| 黄花落叶松林 Larix olgensis forest | 0.47 (55.42) | 0.33 (24.24) | 0.27 (48.15) | 1.07 (0.63) | 50.92 (0.62) | 45.73 (4.25) | 39.69 (5.04) | |
| 人工黄花落叶松林 Larix olgensis plantation | 0.90 (139.25) | 1.56 (33.97) | 0.23 (21.74) | 2.69 (2.57) | 50.82 (0.62) | 45.21 (4.17) | 41.49 (5.50) | |
| 人工红松林 Pinus koraiensis plantation | 0.22 (54.34) | 0.31 (19.35) | 0.56 (21.43) | 1.09 (0.91) | 52.97 (0.43) | 44.36 (3.89) | 36.15 (5.43) | |
| 人工樟子松林 Pinus sylvestris var. mongolica plantation | 0.77 (114.19) | 1.17 (31.62) | 0.38 (49.63) | 2.32 (2.95) | 50.77 (0.80) | 41.82 (1.57) | 35.96 (11.40) | |
| 人工杨树林 Populus plantation | 0.00 (0.00) | 0.00 (0.00) | 0.04 (2.81) | 0.04 (0.05) | 47.61 (0.41) | 0.00 (0.00) | 42.20 (4.14) | |
表1 不同林分类型的林下植被生物量及植被含碳率
Table 1 Biomass of understory vegetation and carbon content of vegetation in different forest types
| 林分类型 Forest type | 林下植被生物量 Biomass of understory vegetation (t·hm-2) | 植被含碳率 Carbon content of vegetation (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 幼树 Young tree | 灌木 Shrub | 草本 Herb | 合计 Total | 乔木 Tree | 灌木 Shrub | 草本 Herb | ||
| 白桦林 Betula platyphylla forest | 1.07 (56.57) | 0.28 (10.71) | 0.63 (34.92) | 1.98 (1.27) | 48.22 (0.70) | 43.58 (4.92) | 40.76 (5.86) | |
| 山杨林 Populus davidiana forest | 0.52 (17.38) | 0.47 (17.02) | 1.34 (35.07) | 2.33 (1.24) | 48.45 (0.55) | 47.25 (1.13) | 42.16 (2.88) | |
| 胡桃楸林 Juglans mandshurica forest | 0.66 (81.86) | 0.42 (7.14) | 0.22 (22.73) | 1.30 (1.31) | 45.80 (0.74) | 42.58 (7.96) | 39.24 (5.52) | |
| 椴树林 Tilia forest | 0.31 (74.19) | 0.53 (20.75) | 0.99 (7.07) | 1.83 (1.38) | 46.77 (0.31) | 45.76 (5.44) | 41.51 (6.44) | |
| 蒙古栎林 Quercus mongolica forest | 0.47 (111.79) | 2.36 (32.63) | 0.68 (22.06) | 3.51 (3.31) | 48.46 (0.41) | 43.69 (4.35) | 37.24 (5.34) | |
| 杂木林 Undesirable tree forest | 0.25 (18.56) | 1.61 (22.36) | 0.70 (35.71) | 2.56 (2.87) | 46.36 (0.43) | 39.79 (7.74) | 42.66 (4.52) | |
| 阔叶混交林 Broadleaf mixed forest | 0.72 (83.30) | 2.26 (22.57) | 0.86 (12.79) | 3.84 (2.59) | 47.60 (2.43) | 43.02 (8.48) | 37.80 (6.88) | |
| 针阔混交林 Needle broad-leaved mixed forest | 0.99 (74.17) | 2.20 (15.00) | 1.26 (17.46) | 4.45 (2.22) | 49.51 (1.85) | 45.44 (6.00) | 40.68 (7.30) | |
| 针叶混交林 Needle mixed forest | 0.96 (111.88) | 0.47 (17.02) | 0.80 (13.75) | 2.23 (1.29) | 51.08 (1.08) | 45.67 (3.32) | 42.62 (2.53) | |
| 黄花落叶松林 Larix olgensis forest | 0.47 (55.42) | 0.33 (24.24) | 0.27 (48.15) | 1.07 (0.63) | 50.92 (0.62) | 45.73 (4.25) | 39.69 (5.04) | |
| 人工黄花落叶松林 Larix olgensis plantation | 0.90 (139.25) | 1.56 (33.97) | 0.23 (21.74) | 2.69 (2.57) | 50.82 (0.62) | 45.21 (4.17) | 41.49 (5.50) | |
| 人工红松林 Pinus koraiensis plantation | 0.22 (54.34) | 0.31 (19.35) | 0.56 (21.43) | 1.09 (0.91) | 52.97 (0.43) | 44.36 (3.89) | 36.15 (5.43) | |
| 人工樟子松林 Pinus sylvestris var. mongolica plantation | 0.77 (114.19) | 1.17 (31.62) | 0.38 (49.63) | 2.32 (2.95) | 50.77 (0.80) | 41.82 (1.57) | 35.96 (11.40) | |
| 人工杨树林 Populus plantation | 0.00 (0.00) | 0.00 (0.00) | 0.04 (2.81) | 0.04 (0.05) | 47.61 (0.41) | 0.00 (0.00) | 42.20 (4.14) | |
| 林分类型 Forest type | 面积 Area (× 104 hm2) | 碳储量 Carbon storage (Tg C) | 碳密度 Carbon density (t·hm-2) | 固碳速率 Carbon sequestration rate (t·hm-2·a-1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2009 | 2014 | 2009 | 2014 | 变化 Change | 2009 | 2014 | 变化 Change | |||||
| 阔叶混交林 Broadleaf mixed forest | 351.22 | 353.56 | 240.65 | 254.34 | 13.69 | 68.52 | 71.93 | 3.41 | 0.75 | |||
| 蒙古栎林 Quercus mongolica forest | 99.80 | 100.44 | 61.07 | 78.52 | 17.45 | 61.19 | 78.15 | 16.96 | 3.48 | |||
| 针阔混交林 Needle broad-leaved mixed forest | 76.34 | 83.38 | 65.86 | 65.97 | 0.11 | 86.26 | 79.07 | -7.19 | 0.03 | |||
| 黄花落叶松林 Larix olgensis forest | 56.94 | 57.58 | 24.97 | 23.52 | -1.45 | 43.85 | 40.81 | -3.04 | -0.51 | |||
| 杨树林 Populus forest | 47.98 | 52.03 | 16.41 | 18.31 | 1.90 | 34.21 | 35.15 | 0.94 | 0.75 | |||
| 针叶混交林 Needle mixed forest | 24.95 | 24.95 | 23.76 | 21.16 | -2.60 | 95.23 | 84.79 | -10.44 | -2.08 | |||
| 杂木林 Undesirable tree forest | 15.78 | 22.18 | 4.09 | 6.64 | 2.55 | 25.86 | 29.95 | 4.09 | 2.60 | |||
| 樟子松林 Pinus sylvestris var. mongolica forest | 12.79 | 11.52 | 5.72 | 3.68 | -2.04 | 47.38 | 34.57 | -12.81 | -3.39 | |||
| 白桦林 Betula platyphylla forest | 11.94 | 11.30 | 6.11 | 6.07 | -0.04 | 51.12 | 53.71 | 2.59 | -0.07 | |||
| 胡桃楸林 Juglans mandshurica forest | 7.89 | 13.01 | 3.50 | 6.81 | 3.31 | 44.47 | 52.37 | 7.90 | 6.04 | |||
| 椴树林 Tilia forest | 5.97 | 8.10 | 4.73 | 6.98 | 2.25 | 79.23 | 86.17 | 6.94 | 6.21 | |||
| 榆树林 Ulmus forest | 5.33 | 3.63 | 2.10 | 1.55 | -0.55 | 39.26 | 42.86 | 3.60 | -2.55 | |||
| 云杉林 Picea forest | 4.48 | 5.12 | 5.48 | 3.85 | -1.63 | 122.28 | 75.30 | -46.98 | -6.70 | |||
| 红松林 Pinus koraiensis forest | 4.05 | 6.40 | 3.90 | 4.77 | 0.87 | 96.14 | 74.49 | -21.65 | 3.19 | |||
| 硕桦林 Betula costata forest | 1.92 | 2.77 | 1.57 | 2.01 | 0.44 | 81.27 | 72.64 | -8.63 | 3.62 | |||
| 水曲柳林 Fraxinus mandshurica forest | 1.49 | 1.07 | 0.60 | 0.67 | 0.07 | 40.27 | 62.43 | 22.16 | 1.13 | |||
| 臭冷杉林 Abies nephrolepis forest | 0.85 | 1.07 | 0.78 | 0.90 | 0.12 | 91.30 | 84.17 | -7.13 | 2.44 | |||
| 黄檗林 Phellodendron amurense forest | 0.00 | 0.21 | 0.00 | 0.01 | 0.01 | 0.00 | 4.51 | 4.51 | 1.59 | |||
| 总计 Total | 729.73 | 758.09 | 471.29 | 505.76 | 34.47 | 64.58 | 66.68 | 2.10 | 0.92 | |||
表2 吉林省森林植被2009年和2014年碳储量、碳密度及其变化
Table 2 Carbon storage and carbon density of forest vegetation in 2009 and 2014 and their changes between 2009 and 2014 in Jilin Province
| 林分类型 Forest type | 面积 Area (× 104 hm2) | 碳储量 Carbon storage (Tg C) | 碳密度 Carbon density (t·hm-2) | 固碳速率 Carbon sequestration rate (t·hm-2·a-1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2009 | 2014 | 2009 | 2014 | 变化 Change | 2009 | 2014 | 变化 Change | |||||
| 阔叶混交林 Broadleaf mixed forest | 351.22 | 353.56 | 240.65 | 254.34 | 13.69 | 68.52 | 71.93 | 3.41 | 0.75 | |||
| 蒙古栎林 Quercus mongolica forest | 99.80 | 100.44 | 61.07 | 78.52 | 17.45 | 61.19 | 78.15 | 16.96 | 3.48 | |||
| 针阔混交林 Needle broad-leaved mixed forest | 76.34 | 83.38 | 65.86 | 65.97 | 0.11 | 86.26 | 79.07 | -7.19 | 0.03 | |||
| 黄花落叶松林 Larix olgensis forest | 56.94 | 57.58 | 24.97 | 23.52 | -1.45 | 43.85 | 40.81 | -3.04 | -0.51 | |||
| 杨树林 Populus forest | 47.98 | 52.03 | 16.41 | 18.31 | 1.90 | 34.21 | 35.15 | 0.94 | 0.75 | |||
| 针叶混交林 Needle mixed forest | 24.95 | 24.95 | 23.76 | 21.16 | -2.60 | 95.23 | 84.79 | -10.44 | -2.08 | |||
| 杂木林 Undesirable tree forest | 15.78 | 22.18 | 4.09 | 6.64 | 2.55 | 25.86 | 29.95 | 4.09 | 2.60 | |||
| 樟子松林 Pinus sylvestris var. mongolica forest | 12.79 | 11.52 | 5.72 | 3.68 | -2.04 | 47.38 | 34.57 | -12.81 | -3.39 | |||
| 白桦林 Betula platyphylla forest | 11.94 | 11.30 | 6.11 | 6.07 | -0.04 | 51.12 | 53.71 | 2.59 | -0.07 | |||
| 胡桃楸林 Juglans mandshurica forest | 7.89 | 13.01 | 3.50 | 6.81 | 3.31 | 44.47 | 52.37 | 7.90 | 6.04 | |||
| 椴树林 Tilia forest | 5.97 | 8.10 | 4.73 | 6.98 | 2.25 | 79.23 | 86.17 | 6.94 | 6.21 | |||
| 榆树林 Ulmus forest | 5.33 | 3.63 | 2.10 | 1.55 | -0.55 | 39.26 | 42.86 | 3.60 | -2.55 | |||
| 云杉林 Picea forest | 4.48 | 5.12 | 5.48 | 3.85 | -1.63 | 122.28 | 75.30 | -46.98 | -6.70 | |||
| 红松林 Pinus koraiensis forest | 4.05 | 6.40 | 3.90 | 4.77 | 0.87 | 96.14 | 74.49 | -21.65 | 3.19 | |||
| 硕桦林 Betula costata forest | 1.92 | 2.77 | 1.57 | 2.01 | 0.44 | 81.27 | 72.64 | -8.63 | 3.62 | |||
| 水曲柳林 Fraxinus mandshurica forest | 1.49 | 1.07 | 0.60 | 0.67 | 0.07 | 40.27 | 62.43 | 22.16 | 1.13 | |||
| 臭冷杉林 Abies nephrolepis forest | 0.85 | 1.07 | 0.78 | 0.90 | 0.12 | 91.30 | 84.17 | -7.13 | 2.44 | |||
| 黄檗林 Phellodendron amurense forest | 0.00 | 0.21 | 0.00 | 0.01 | 0.01 | 0.00 | 4.51 | 4.51 | 1.59 | |||
| 总计 Total | 729.73 | 758.09 | 471.29 | 505.76 | 34.47 | 64.58 | 66.68 | 2.10 | 0.92 | |||

图1 2009年和2014年吉林省森林植被不同龄组碳储量(A)和碳密度(B)。I, 幼龄林; II, 中龄林; III, 近熟林; IV, 成熟林; V, 过熟林。
Fig. 1 Carbon storage (A) and density (B) of forest vegetation in different age classes in Jilin Province in 2009 and 2014. I, young forest; II, middle- aged forest; III, near mature forest; IV, mature forest; V, over mature forest.
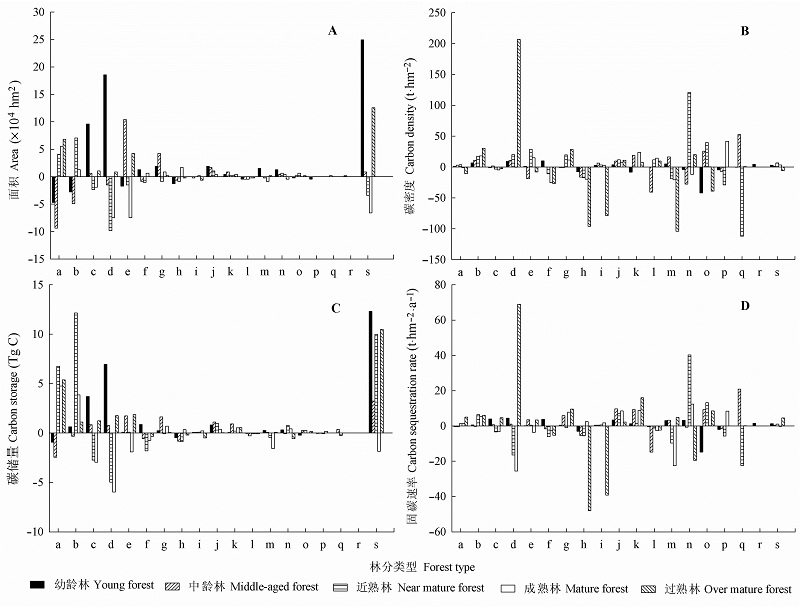
图2 2009年和2014年吉林省森林植被不同林分类型和龄组林分面积(A)、碳密度(B)、碳储量(C)和固碳速率(D)的变化。a, 阔叶混交林; b, 蒙古栎林; c, 针阔混交林; d, 黄花落叶松林; e, 杨树林; f, 针叶混交林; g, 杂木林; h, 樟子松林; i, 白桦林; j, 胡桃楸林; k, 椴树林; l, 榆树林; m, 云杉林; n, 红松林; o, 硕桦林; p, 水曲柳林; q, 臭冷杉林; r, 黄檗林; s, 总计。
Fig. 2 Changes of forest area (A), carbon density (B), carbon storage (C), and carbon sequestration rate (D) of different forest types and age classes in Jilin Province in 2009 and 2014. a, broadleaf mixed forest; b, Quercus mongolica forest; c, needle broad-leaved mixed forest; d, Larix olgensis forest; e, Populus forest; f, needle mixed forest; g, undesirable tree forest; h, Pinus sylvestris var. mongolica forest; i, Betula platyphylla forest; j, Juglans mandshurica forest; k, Tilia forest; l, Ulmus forest; m, Picea forest; n, Pinus koraiensis forest; o, Betula costata forest; p, Fraxinus mandshurica forest; q, Abies nephrolepis forest; r, Phellodendron amurense forest; s, total.
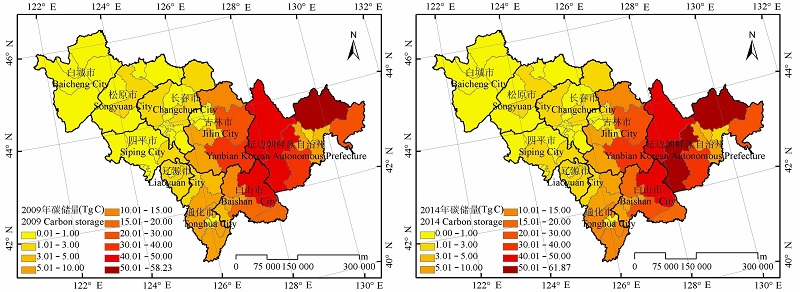
图3 吉林省森林植被碳储量的分布。
Fig. 3 Distribution of forest vegetation carbon storage in Jilin Province.
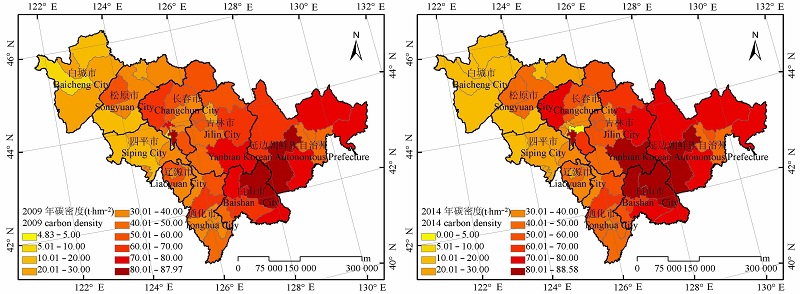
图4 吉林省森林植被碳密度的分布。
Fig. 4 Distribution of forest vegetation carbon density in Jilin Province.
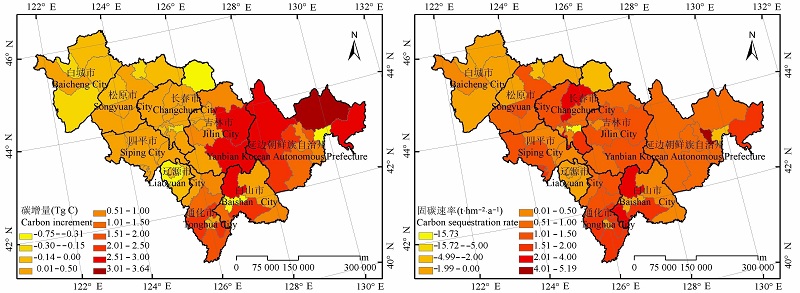
图5 森林植被碳增量与固碳速率的分布。
Fig. 5 Distribution of carbon increment and carbon sequestration rate of forest vegetation in Jilin Province.
| 1 | Baccini A, Goetz SJ, Walker WS, Laporte NT, Sun M, Sulla-Menashe D, Hackler J, Beck PSA, Dubayah R, Friedl MA, Samanta S, Houghton RA (2012). Estimated carbon dioxide emissions from tropical deforestation improved by carbon-density maps.Nature Climate Change, 2, 182-185. |
| 2 | Chastain Jr RA, Currie WS, Townsend PA (2006). Carbon sequestration and nutrient cycling implications of the evergreen understory layer in Appalachian forests.Forest Ecology and Management, 231, 63-77. |
| 3 | Chen CG, Zhu JF (1989). The Manual for Main Tree Biomass in Northeast China. China Forestry Publishing House, Beijing. 158-173. (in Chinese)[陈传国, 朱俊凤(1989). 东北主要林木生物量手册. 中国林业出版社, 北京. 158-173.] |
| 4 | Chen DK, Zhou XF, Ding BY, Hu ZC, Zhu N, Wang YH, Zhao HX, Ju YG, Jin YY (1984). Research on natural secondary forest in Heilongjing Province—The Management approach of planting conifers and conserving deciduous trees.Journal of North-Eastern Forestry Institute, 12(4), 1-12. (in Chinese with English abstract)[陈大珂, 周晓峰, 丁宝永, 胡正昌, 祝宁, 王义弘, 赵惠勋, 鞠永贵, 金永岩 (1984). 黑龙江省天然次生林研究(I)——栽针保阔的经营途径. 东北林学院学报, 12(4), 1-12.] |
| 5 | Chen XL (2003). Researches on Carbon Sequestration Functions of Main Forest Types in Northern China. PhD dissertation, Beijing Forestry University, Beijing. 84-85. (in Chinese with English abstract) |
| 6 | [陈遐林 (2003). 华北主要森林类型的碳汇功能研究. 博士学位论文, 北京林业大学, 北京. 84-85.] |
| 7 | Dixon RK, Brown S, Houghton RA, Solomon AM, Trexler MC, Wisniewski J (1994). Carbon pools and flux of global forest ecosystems.Science, 263, 185-190. |
| 8 | Fan CN (2014). The Carbon Distribution Characteristic and Storage of Forest Vegetation in Jilin Province. PhD dissertation, Northeast Forestry University, Harbin. 27-33. (in Chinese with English abstract)[范春楠 (2014). 吉林省森林植被碳估算及其分布特征. 博士学位论文, 东北林业大学, 哈尔滨. 27-33.] |
| 9 | Fan CN, Pang SJ, Zheng JP, Li B, Guo ZL (2013). Biomass estimating models of saplings for 14 species in Changbaishan Mountains, Northeastern China.Journal of Beijing Forestry University, 35(2), 1-9. (in Chinese with English abstract)[范春楠, 庞圣江, 郑金萍, 李兵, 郭忠玲 (2013). 长白山林区14种幼树生物量估测模型. 北京林业大学学报, 35(2), 1-9.] |
| 10 | Fang JY, Chen AP, Peng CH, Zhao SQ, Ci LJ (2001). Changes in forest biomass carbon storage in China between 1949 and 1998.Science, 292, 2320-2322. |
| 11 | Gilliam FS (2007). The ecological significance of the herbaceous layer in temperate forest ecosystems.Bioscience, 57, 845-858. |
| 12 | Hu HQ, Luo BZ, Wei SJ, Wei SW, Sun L, Luo SS, Ma HB (2015). Biomass carbon density and carbon sequestration capacity in seven typical forest types of the Xiaoxing’an Mountains, China.Chinese Journal of Plant Ecology, 39, 140-158. (in Chinese with English abstract)[胡海清, 罗碧珍, 魏书精, 魏书威, 孙龙, 罗斯生, 马洪斌 (2015). 小兴安岭7种典型林型林分生物量碳密度与固碳能力. 植物生态学报, 39, 140-158.] |
| 13 | Hu Q, Wang SL, Chen LC, Zhang WD, Gong C (2012). Biomass and carbon density of main forest ecosystems in Hubei Province.Chinese Journal of Ecology, 31, 1626-1632. (in Chinese with English abstract)[胡青, 汪思龙, 陈龙池, 张伟东, 宫超 (2012). 湖北省主要森林类型生态系统生物量与碳密度比较. 生态学杂志, 31, 1626-1632.] |
| 14 | IPCC (Intergovernmental Panel on Climate Change) (2006). Agriculture, forestry and other land use. In: Eggleston HS, Buendia L, Miwa K, Ngara T, Tanabe K eds. 2006 IPCC Guidelines for National Greenhouse Gas Inventories. Institute for Global Environmental Strategies, Kanagawa, Japan. |
| 15 | Jia WW, Jiang SW, Li FR (2008). Biomass of single tree of Pinus sylvestris var. mongolica in Eastern Heilongjiang.Liaoning Forestry Science and Technology, (3), 5-9. (in Chinese with English abstract)[贾炜玮, 姜生伟, 李凤日 (2008). 黑龙江东部地区樟子松人工林单木生物量研究. 辽宁林业科技, (3), 5-9.] |
| 16 | Jiao Y, Hu HQ (2005). Carbon storage and its dynamics of forest vegetations in Heilongjiang Province.Chinese Journal of Applied Ecology, 16, 2248-2252. (in Chinese with English abstract)[焦燕, 胡海清 (2005). 黑龙江省森林植被碳储量及其动态变化. 应用生态学报, 16, 2248-2252.] |
| 17 | Keeling CD, Chin JFS, Whorf TP (1996). Increased activity of northern vegetation inferred from atmospheric CO2 measurements.Nature, 382, 146-149. |
| 18 | Lamlom SH, Savidge RA (2003). A reassessment of carbon content in wood: Variation within and between 41 North American species.Biomass and Bioenergy, 25, 381-388. |
| 19 | Li HK, Zhao PX, Lei YC, Zeng WS (2012). Comparison on estimation of wood biomass using forest inventory data.Scientia Silvae Sinicae, 48(5), 44-52. (in Chinese with English abstract)[李海奎, 赵鹏祥, 雷渊才, 曾伟生 (2012). 基于森林清查资料的乔木林生物量估算方法的比较. 林业科学, 48(5), 44-52.] |
| 20 | Li KR, Wang SQ, Cao MK (2004). Vegetation and soil carbon storage in China. Science in China (Series D), 33(1), 72-80. (in Chinese)[李克让, 王绍强, 曹明奎 (2003). 中国植被和土壤碳贮量. 中国科学(D辑), 33(1), 72-80.] |
| 21 | Liu GH, Fu BJ, Fang JY (2000). Carbon dynamics of Chinese forests and its contribution to global carbon balance.Acta Ecologica Sinica, 20, 733-740. (in Chinese with English abstract)[刘国华, 傅伯杰, 方精云 (2000). 中国森林碳动态及其对全球碳平衡的贡献. 生态学报, 20, 733-740.] |
| 22 | Luan JW, Liu SR, Zhu XL, Wang JX (2011). Soil carbon stocks and fluxes in a warm-temperate oak chronosequence in China.Plant and Soil, 347, 243-253. |
| 23 | Mu CC, Zhuang C, Han YR, Zhang XL, Tong JQ, Cheng JY, Cao WL (2014). Effect of liberation cutting on the vegetation carbon storage of Korean pine forests by planting conifer and reserving broad-leaved tree in Changbai Mountains of China.Bulletin of Botanical Research, 34, 529-536. (in Chinese with English abstract)[牟长城, 庄宸, 韩阳瑞, 张晓亮, 佟静秋, 程家友, 曹万亮 (2014). 透光抚育对长白山“栽针保阔”红松林植被碳储量影响. 植物研究, 34,529-536.] |
| 24 | Nowak DJ, Crane DE (2002). Carbon storage and sequestration by urban trees in the USA.Environmental Pollution, 116, 381-389. |
| 25 | Qi L, Yu DP, Zhou WM, Zhou L, Zhao FA, Wang CH, Dai LM (2013). Impact of logging on carbon density of broadleaved-Korean pine mixed forests on Changbai Mountains.Acta Ecologica Sinica, 33, 3065-3073. (in Chinese with English abstract)[齐麟, 于大炮, 周旺明, 周莉, 赵福安, 王长海, 代力民 (2013). 采伐对长白山阔叶红松林生态系统碳密度的影响. 生态学报, 33, 3065-3073.] |
| 26 | Schlesinger WH (1977). Carbon balance in terrestrial detritus.Annual Review of Ecology and Systematics, 8, 51-81. |
| 27 | Schroeder P, Brown S, Mo J, Birdsey R, Cleszewski C (1997). Biomass estimation for temperate broadleaf forests of the United States using inventory data.Forest Science, 43, 424-434. |
| 28 | Tang FD, Han SJ, Zhang JH (2009). Carbon dynamics of broad-leaved Korean pine forest ecosystem in Changbai Mountains and its responses to climate change.Chinese Journal of Applied Ecology, 20, 1285-1292. (in Chinese with English abstract)[唐凤德, 韩士杰, 张军辉 (2009). 长白山阔叶红松林生态系统碳动态及其对气候变化的响应. 应用生态学报, 20, 1285-1292.] |
| 29 | Tans PP, Fung IY, Takahashi T (1990). Observational contrains on the global atmospheric CO2 budget.Science, 247, 1431-1438. |
| 30 | Taylor AH, Huang JY, Zhou SQ (2004). Canopy tree development and undergrowth bamboo dynamics in old-growth Abies-Betula forests in southwestern China: A 12-year study.Forest Ecology and Management, 200, 347-360. |
| 31 | Tian YY, Qin F, Yan H, Guo WH, Guan QW (2011). Carbon content rate in the common woody plants of China.Journal of Anhui Agricultural Sciences, 39, 16166-16169. (in Chinese with English abstract)[田勇燕, 秦飞, 言华, 郭伟红, 关庆伟 (2011). 我国常见木本植物的含碳率. 安徽农业科学, 39, 16166-16169.] |
| 32 | Wang CK (2006). Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests.Forest Ecology and Management, 222, 9-16. |
| 33 | Wang JL, Wang XH, Yue CR, Cheng F, Xu TS, Cheng PF, Wang XM, Gao Y (2012). Carbon content rate in dominant species of four forest types in Shangri-la, northwest Yunnan Province.Ecology and Environment Sciences, 21, 613-619. (in Chinese with English abstract)[王金亮, 王小花, 岳彩荣, 程峰, 徐天蜀, 程鹏飞, 王雪梅, 高雁 (2012). 滇西北香格里拉森林4个建群种的含碳率. 生态环境学报, 21, 613-619.] |
| 34 | Wang RH, Ge XM, Tang LZ (2014). A review of diversity, biomass and nutrient effect of understory vegetation.World Forestry Research, 27, 43-48. (in Chinese with English abstract)[王瑞华, 葛晓敏, 唐罗忠 (2014). 林下植被多样性、生物量及养分作用研究进展. 世界林业研究, 27, 43-48.] |
| 35 | Wang SQ, Liu JY, Yu GR (2003). Error analysis of estimating terrestrial soil organic carbon storage in China.Chinese Journal of Applied Ecology, 14, 797-802. (in Chinese with English abstract)[王绍强, 刘纪远, 于贵瑞 (2003). 中国陆地土壤有机碳蓄积量估算误差分析. 应用生态学报, 14, 797-802.] |
| 36 | Wang XC, Qi G, Yu DP, Zhou L, Dai LM (2011). Carbon storage, density, and distribution in forest ecosystems in Jilin Province of Northeast China.Chinese Journal of Applied Ecology, 22, 2013-2020. (in Chinese with English abstract)[王新闯, 齐光, 于大炮, 周莉, 代力民 (2011). 吉林省森林生态系统的碳储量、碳密度及其分布. 应用生态学报, 22, 2013-2020.] |
| 37 | Wang XK, Bai YY, Ouyang ZY, Miao H (2002). Missing sink in global carbon cycle and its causes.Acta Ecologica Sinica, 22, 94-103. (in Chinese with English abstract)[王效科, 白艳莹, 欧阳志云, 苗鸿 (2002). 全球碳循环中的失汇及其形成原因. 生态学报, 22, 94-103.] |
| 38 | Wang XK, Feng ZW, Ouyang ZY (2001). Vegetation carbon storage and density of forest ecosystems in China.Chinese Journal of Applied Ecology, 12, 13-16. (in Chinese with English abstract)[王效科, 冯宗炜, 欧阳志云 (2001). 中国森林生态系统的植物碳储量和碳密度研究. 应用生态学报, 12, 13-16.] |
| 39 | Wani AA, Joshi PK, Singh O, Bhat JA (2014). Estimating soil carbon storage and mitigation under temperate coniferous forests in the southern region of Kashmir Himalayas.Mitigation and Adaptation Strategies for Global Change, 19, 1179-1194. |
| 40 | Xie XL, Sun B, Zhou HZ, Li ZP, Li AB (2004). Organic carbon density and storage in soils of china and spatial analysis.Acta Pedologica Sinica, 41, 35-43. (in Chinese with English abstract)[解宪丽, 孙波, 周慧珍, 李忠佩, 李安波 (2004). 中国土壤有机碳密度和储量的估算与空间分布分析. 土壤学报, 41, 35-43.] |
| 41 | Yamamoto SI, Nishimura N, Matsui K (1995). Natural disturbance and tree species coexistence in an old-growth beech-dwarf bamboo forest, southwestern Japan.Journal of Vegetation Science, 6, 875-886. |
| 42 | Yang C, Tian DL, Hu YL, Yan WD, Fang X, Liang XC (2011). Dynamics of understory vegetation biomass in successive |
| 43 | rotations of Chinese fir (Cunninghamia lanceolata) plantations.Acta Ecologica Sinica, 31, 2737-2447. (in Chinese with English abstract)[杨超, 田大伦, 胡曰利, 闫文德, 方晰, 梁小翠 (2011). 连栽杉木林林下植被生物量动态格局. 生态学报, 31, 2737-2447.] |
| 44 | Yang K, Guan DS (2007). Selection of gaining quadrat for harvesting the undergrowth vegetation and its biomass estimation modeling in forest.Acta Ecologica Sinica, 27, 705-714.(in Chinese with English abstract) [杨昆, 管东生 (2007). 森林林下植被生物量收获的样方选择和模型. 生态学报, 27, 705-714.] |
| 45 | Yu Y, Fan WY, Li MZ (2012). Forest carbon rates at different scales in Northeast China forest area.Chinese Journal of Applied Ecology, 23, 341-346. (in Chinese with English abstract)[于颖, 范文义, 李明泽 (2012). 东北林区不同尺度森林的含碳率. 应用生态学报, 23, 341-346.] |
| 46 | Zhao M, Zhou GS (2004). Carbon storage of forest vegetation and its relationship with climatic factors.Scientia Geographica Sinica, 24, 50-54. (in Chinese with English abstract)[赵敏, 周广胜 (2004). 中国森林生态系统的植物碳贮量及其影响因子分析. 地理科学, 24, 50-54.] |
| 47 | Zhen W, Huang M, Zhai YL, Chen K, Gong YZ (2014). Variation of forest vegetation carbon storage and carbon sequestration rate in Liaoning Province, Northeast China.Chinese Journal of Applied Ecology, 25, 1259-1265. (in Chinese with English abstract)[甄伟, 黄玫, 翟印礼, 陈珂, 龚亚珍 (2014). 辽宁省森林植被碳储量和固碳速率变化. 应用生态学报, 25, 1259-1265.] |
| 48 | Zheng ZM, Zhang M, Wen XF, Sun XM, Yu GR, Zhang LM, Han SJ, Wu JB (2009). Contributions of below-canopy CO2 fluxes to ecosystem carbon budget of a temperate mixed forest in Changbai Mountain.Acta Ecologica Sinica, 29, 1-8. (in Chinese with English abstract)[郑泽梅, 张弥, 温学发, 孙晓敏, 于贵瑞, 张雷明, 韩士杰, 吴家兵 (2009). 长白山温带混交林林冠下层CO2通量对生态系统碳收支的贡献. 生态学报, 29, 1-8.] |
| 49 | Zhou GY, Liu SG, Li ZA, Zhang DQ, Tang XL, Zhou CY, Yan JH, Mo JM (2006). Old-growth forests can accumulate carbon in soils.Science, 314, 1417. |
| 50 | Zhou YR, Yu ZL, Zhao SD (2000). Carbon storage and budget of major Chinese forest types.Acta Phytoecologica Sinica, 24, 518-522. (in Chinese with English abstract)[周玉荣, 于振良, 赵士洞 (2000). 我国主要森林生态系统碳贮量和碳平衡. 植物生态学报, 24, 518-522.] |
| 51 | Zhu B, Wang XP, Fang JY, Piao SL, Shen HH, Zhao SQ, Peng CH (2010). Altitudinal changes in carbon storage of temperate forests on Mt. Changbai, Northeast China.Journal of Plant Research, 123, 439-452. |
| [1] | 张智洋 赵颖慧 甄贞. 1986-2022年松花江流域陆地生态系统碳储量动态监测[J]. 植物生态学报, 2024, 48(预发表): 0-0. |
| [2] | 张计深, 史新杰, 刘宇诺, 吴阳, 彭守璋. 气候变化下中国潜在自然植被生态系统碳储量动态[J]. 植物生态学报, 2024, 48(4): 428-444. |
| [3] | 李晓田, 王铁娟, 韩文娟, 张丽, 张慧, 刘晓婷, 刘雅洁. 东阿拉善珍稀濒危植物绵刺种群结构与点格局分析[J]. 植物生态学报, 2023, 47(4): 506-514. |
| [4] | 石荡, 郭传超, 蒋南林, 唐莹莹, 郑凤, 王瑾, 廖康, 刘立强. 新疆野杏天然更新幼株的个体特征及空间分布格局[J]. 植物生态学报, 2023, 47(4): 515-529. |
| [5] | 徐干君, 吴胜义, 李伟, 赵欣胜, 聂磊超, 唐希颖, 翟夏杰. 陕西黄河湿地自然保护区碳储量估算[J]. 植物生态学报, 2023, 47(4): 469-478. |
| [6] | 闫涵, 马松梅, 魏博, 张宏祥, 张丹. 孑遗灌木长柄扁桃的历史分布格局及其环境驱动力[J]. 植物生态学报, 2022, 46(7): 766-774. |
| [7] | 张央, 安明态, 武建勇, 刘锋, 汪伟. 中国兜兰属宽瓣亚属植物地理分布格局及其主导气候因子[J]. 植物生态学报, 2022, 46(1): 40-50. |
| [8] | 左永令, 杨小波, 李东海, 吴二焕, 杨宁, 李龙, 张培春, 陈琳, 李晨笛. 环境因子对海南岛野生兰科植物物种组成与分布格局的影响[J]. 植物生态学报, 2021, 45(12): 1341-1349. |
| [9] | 丁凯, 张毓婷, 张俊红, 柴雄, 周世水, 童再康. 不同密度杉木林对林下植被和土壤微生物群落结构的影响[J]. 植物生态学报, 2021, 45(1): 62-73. |
| [10] | 张亮, 王志磊, 薛婷婷, 郝笑云, 杨晨露, 高飞飞, 王莹, 韩星, 李华, 王华. 葡萄园生态系统碳源/汇及碳减排策略研究进展[J]. 植物生态学报, 2020, 44(3): 179-191. |
| [11] | 扈明媛, 袁野, 戴晓琴, 付晓莉, 寇亮, 王辉民. 亚热带人工林乔灌草根际土壤氮矿化特征[J]. 植物生态学报, 2020, 44(12): 1285-1295. |
| [12] | 拓锋, 刘贤德, 刘润红, 赵维俊, 敬文茂, 马剑, 武秀荣, 赵晶忠, 马雪娥. 祁连山大野口流域青海云杉种群空间格局及其关联性[J]. 植物生态学报, 2020, 44(11): 1172-1183. |
| [13] | 唐丽丽, 杨彤, 刘鸿雁, 康慕谊, 王仁卿, 张峰, 高贤明, 岳明, 张梅, 郑璞帆, 石福臣. 华北地区荆条灌丛分布及物种多样性空间分异 规律[J]. 植物生态学报, 2019, 43(9): 825-833. |
| [14] | 吴盼, 彭希强, 杨树仁, 高亚男, 白丰桦, 衣世杰, 杜宁, 郭卫华. 山东省滨海湿地柽柳种群的空间分布格局及其关联性[J]. 植物生态学报, 2019, 43(9): 817-824. |
| [15] | 蔡琴, 丁俊祥, 张子良, 胡君, 汪其同, 尹明珍, 刘庆, 尹华军. 青藏高原东缘主要针叶树种叶片碳氮磷化学计量分布格局及其驱动因素[J]. 植物生态学报, 2019, 43(12): 1048-1060. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19