
植物生态学报 ›› 2017, Vol. 41 ›› Issue (11): 1127-1139.DOI: 10.17521/cjpe.2017.0092
• 研究论文 • 下一篇
马明哲1,2, 申国珍1, 熊高明1, 赵常明1, 徐文婷1, 周友兵1, 谢宗强1,2,*( )
)
收稿日期:2017-04-07
接受日期:2017-08-29
出版日期:2017-11-10
发布日期:2017-11-10
通讯作者:
谢宗强
基金资助:
Ming-Zhe MA1,2, Guo-Zhen SHEN1, Gao-Ming XIONG1, Chang-Ming ZHAO1, Wen-Ting XU1, You-Bing ZHOU1, Zong-Qiang XIE1,2,*()
Received:2017-04-07
Accepted:2017-08-29
Online:2017-11-10
Published:2017-11-10
Contact:
Zong-Qiang XIE
摘要:
充分认识并掌握我国自然遗产地山地植被垂直带谱代表性, 对正确评估自然遗产地的价值进而制定相关保护管理政策具有重要的科学意义和现实意义。该研究基于群落调查数据、全球1 km2土地利用数据, 通过对比分析、空间分析等方法, 从植被垂直带谱的地带性、完整性及不同垂直带群落物种更替等角度, 分析论证了神农架自然遗产地植被垂直带谱的代表性。结果显示: 神农架自然遗产地从低海拔到高海拔依次发育有常绿阔叶林带(遗产地南坡)、常绿落叶阔叶混交林带、落叶阔叶林带、针阔混交林带、针叶林带及亚高山灌丛和草甸带, 其北坡保存的地带性常绿落叶阔叶混交林是北半球常绿落叶阔叶混交林生态系统的最典型代表。神农架自然遗产地拥有的植被垂直带谱是“全球生物地理区划”中东方落叶林生物地理省最完整的植被垂直带谱, 在东方落叶林生物地理省具有唯一性和代表性, 在较小的水平距离范围内浓缩了中亚热带、北亚热带、暖温带、温带和寒温带等生态系统特征, 成为研究全球气候变化下山地生态系统垂直分异规律及其生态学过程的杰出范例, 具有突出的世界自然遗产价值。
马明哲, 申国珍, 熊高明, 赵常明, 徐文婷, 周友兵, 谢宗强. 神农架自然遗产地植被垂直带谱的特点和代表性. 植物生态学报, 2017, 41(11): 1127-1139. DOI: 10.17521/cjpe.2017.0092
Ming-Zhe MA, Guo-Zhen SHEN, Gao-Ming XIONG, Chang-Ming ZHAO, Wen-Ting XU, You-Bing ZHOU, Zong-Qiang XIE. Characteristic and representativeness of the vertical vegetation zonation along the altitudinal gradient in Shennongjia Natural Heritage. Chinese Journal of Plant Ecology, 2017, 41(11): 1127-1139. DOI: 10.17521/cjpe.2017.0092
| 国家 Country | 地区 Area | 植被类型 Vegetation type | 位置 Location | 海拔 Altitude (m) | 年平均气温 Mean annual temperature (℃) | 年降水量 Mean annual precipitation (mm) | 优势种 Dominant species | 土壤类型 Soil type |
|---|---|---|---|---|---|---|---|---|
| 中国 China | 神农架 Shennongjia | 常绿落叶阔叶混交林 Evergreen deciduous broad-leaved mixed forest | 31.27° N, 110.48° E | 1 670 | 10.6 | 1 200 | 米心水青冈, 青冈 Fagus engleriana, Cyclobalanopsis glauca | 山地黄棕壤 Mountain yellow brown soil |
| 日本 Japan | 本州南部低地, 四国, 九州和琉球群岛的北部 The southern lowlands of Honshu, north part of Shikoku, Kyushu and Ryuryu | 常绿阔叶林 Evergreen broad-leaved forest | 26.58°-35.5° N, 130°- 140° E | 250-1 100 | 13-21 | 1 300-2 500 | Castanopsis cuspidata, C. cuspidate var. sieboldii, Quercus salicina, Q. acuta, Q. sessilifolia, Machilus thunbergii | 棕壤 Brown soil |
| 法国、意大利、西班牙、 希腊 France, Italy, Spain, and Greece, | 地中海沿岸 Mediterranean | 硬叶常绿阔叶林 Sclerophyllous evergreen broad-leaved forest | 30.25°- 45.83° N, 0°-30° E | 300-2 800 | 16-20 | 300-1 000 | Quercus coccifera, Q. ilex, Q. suber, Pistacia lentiscus | 红壤和黄壤 Red soil and yellow soil |
| 美国 USA | 大烟山 Great Smoky Mountains | 温带阔叶林 Temperate broad-leaved forest | 35.6° N, 83.48° W | 1 830 | 12 | 1 270-2 030 | Quercus borealis, Tilia heterophylla, Betula alleghenicnsis, Acer rubrum | 山地棕壤 Mountain brown soil |
| 北非 North Africa | 撒哈拉沙漠北部 North of the Sahara Desert | 荒漠 Desert | 25°-35° N, 15°W-50° E | - | >30 | 90-450 | 金合欢属, 蒿属, 三芒草属, 画眉草属, 稷属 Acacia, Artemisia, Aristida, Eragrostis, Panicum | 石漠(岩漠)、砾漠和沙漠 Rock desert (rock desert), gravel desert and desert |
表1 神农架及北半球同纬度典型地区概况
Table 1 Overview of Shennongjia and its counterparts in the same latitude
| 国家 Country | 地区 Area | 植被类型 Vegetation type | 位置 Location | 海拔 Altitude (m) | 年平均气温 Mean annual temperature (℃) | 年降水量 Mean annual precipitation (mm) | 优势种 Dominant species | 土壤类型 Soil type |
|---|---|---|---|---|---|---|---|---|
| 中国 China | 神农架 Shennongjia | 常绿落叶阔叶混交林 Evergreen deciduous broad-leaved mixed forest | 31.27° N, 110.48° E | 1 670 | 10.6 | 1 200 | 米心水青冈, 青冈 Fagus engleriana, Cyclobalanopsis glauca | 山地黄棕壤 Mountain yellow brown soil |
| 日本 Japan | 本州南部低地, 四国, 九州和琉球群岛的北部 The southern lowlands of Honshu, north part of Shikoku, Kyushu and Ryuryu | 常绿阔叶林 Evergreen broad-leaved forest | 26.58°-35.5° N, 130°- 140° E | 250-1 100 | 13-21 | 1 300-2 500 | Castanopsis cuspidata, C. cuspidate var. sieboldii, Quercus salicina, Q. acuta, Q. sessilifolia, Machilus thunbergii | 棕壤 Brown soil |
| 法国、意大利、西班牙、 希腊 France, Italy, Spain, and Greece, | 地中海沿岸 Mediterranean | 硬叶常绿阔叶林 Sclerophyllous evergreen broad-leaved forest | 30.25°- 45.83° N, 0°-30° E | 300-2 800 | 16-20 | 300-1 000 | Quercus coccifera, Q. ilex, Q. suber, Pistacia lentiscus | 红壤和黄壤 Red soil and yellow soil |
| 美国 USA | 大烟山 Great Smoky Mountains | 温带阔叶林 Temperate broad-leaved forest | 35.6° N, 83.48° W | 1 830 | 12 | 1 270-2 030 | Quercus borealis, Tilia heterophylla, Betula alleghenicnsis, Acer rubrum | 山地棕壤 Mountain brown soil |
| 北非 North Africa | 撒哈拉沙漠北部 North of the Sahara Desert | 荒漠 Desert | 25°-35° N, 15°W-50° E | - | >30 | 90-450 | 金合欢属, 蒿属, 三芒草属, 画眉草属, 稷属 Acacia, Artemisia, Aristida, Eragrostis, Panicum | 石漠(岩漠)、砾漠和沙漠 Rock desert (rock desert), gravel desert and desert |
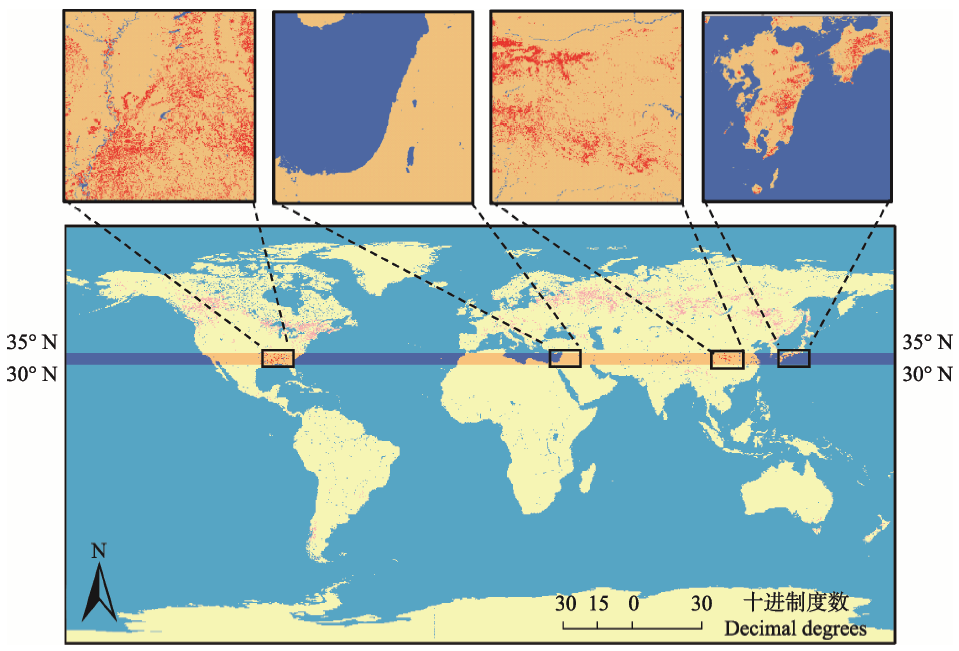
图1 全球30°-35° N带上常绿落叶阔叶混交林的分布(红色代表混交林)。
Fig. 1 The distribution of evergreen deciduous broad-leaved mixed forests (red areas) along the belt of 30°-35° N in the Northern Hemisphere.
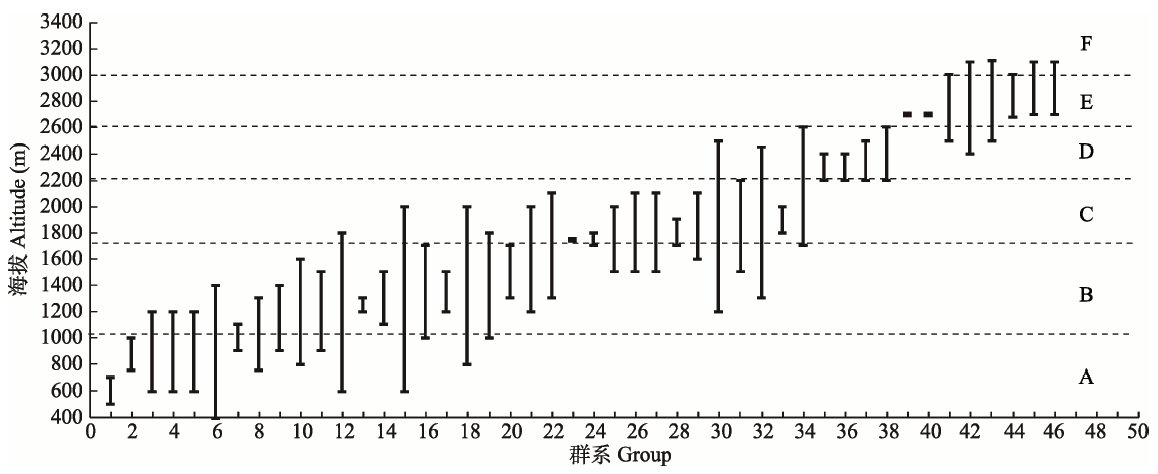
图2 神农架海拔梯度上主要植物群系分布图。大写英文字母对应各垂直带: A, 亚热带常绿阔叶林带; B, 北亚热带常绿落叶阔叶混交林带; C, 暖温带落叶阔叶林带; D, 温带针阔混交林带; E, 寒温带针叶林带; F, 亚高山灌丛、草甸带。 横坐标数字对应群系: 1, 蜡梅灌丛; 2, 楠木、小叶青冈为主的常绿阔叶林; 3, 马桑、毛黄栌灌丛; 4, 马尾松、栓皮栎林; 5, 杉木林; 6, 栓皮栎林; 7, 香叶树、小叶青冈、化香树、亮叶桦林; 8, 马尾松林; 9, 尖齿高山栎灌丛; 10, 曼青冈、水丝梨、巴东栎、青冈林; 11, 乌冈栎、岩栎、鹅耳枥、化香树林; 12, 野核桃林; 13, 栓皮栎、锐齿槲栎、茅栗林; 14, 巴东栎、曼青冈、亮叶桦、化香树林; 15, 刺叶栎林; 16, 短柄枹林; 17, 茅栗林; 18, 巴山松林; 19, 亮叶桦、化香树、鹅耳枥林; 20, 华山松、糙皮桦林; 21, 锐齿槲栎林; 22, 秦岭冷杉林; 23, 川榛、鸡树条荚蒾、湖北海棠灌丛; 24, 薹草、地榆、香青、血见愁老鹳草草甸; 25, 野漆树、锐齿槲栎、灯台树、化香树林; 26, 芒、蕨草丛; 27, 美丽胡枝子、绿叶胡枝子灌丛; 28, 薹草、葱状灯芯草、长叶地榆、柳兰沼泽化草甸; 29, 华山松、锐齿槲栎林; 30, 华山松林; 31, 米心水青冈林; 32, 秦岭冷杉、青扦林; 33, 锐齿槲栎、米心水青冈、红桦林; 34, 红桦林; 35, 华山松、山杨、红桦林; 36, 华山松、山杨林; 37, 中华黄花柳、华中山楂、湖北花楸灌丛; 38, 巴山冷杉、红桦、槭类林; 39, 杯腺柳灌丛; 40, 直穗小檗灌丛; 41, 箭竹灌丛; 42, 平枝荀子灌丛; 43, 巴山冷杉林; 44, 粉红杜鹃灌丛; 45, 香柏灌丛; 46, 印度三毛草、紫羊茅、糙野青茅草甸。群系海拔分布信息主要参考田自强(2002)。
Fig. 2 The distribution of main plant formations in Shennongjia along the elevation gradient. Capital English letters represent vertical vegetation zones: A, subtropical zone of evergreen broad-leaved forest; B, north subtropical zone of Mixed evergreen and deciduous broad-leaved forest; C, warm temperate zone of broadleaved deciduous forest; D, temperate zone of mixed broadleaf-conifer forest; E, cold temperate zone of coniferous forest; F, subalpine zone of shrub meadow. The abscissa represents different formations: 1, Form. Chimonanthus praecox; 2, Form. Phoebe zhennan, Cyclobalanopsis gracilis; 3, Form. Coriaria sinica, Cotinus coggyqria var. pubescens; 4, Form. Pinus massoniana, Quercus variabilis; 5, Form. Cunninghamia lanceolata; 6, Form. Quercus variabilis; 7, Form. Lindera communis, Cyclobalanopsis gracilis, Platycarya strobilacea, Betula luminifera; 8, Form. Pinus massoniana; 9, Form. Quercus acrodonta; 10, Form. Cyclobalanopsis oxyodon, Sycopsis chinense, Quercus engleriana, Cyclobanopsis myrsinaefolia; 11, Form. Quercus philyraeoides, Q. acrodonta, Carpinus sp., Platycarya strobilacea; 12, Form. Juglans cathayensis; 13, Form. Quercus variabilis, Q. aliena var. acuteserrata, Castanea segyinii; 14, Form. Quercus engleriana, Cyclobalanopsis oxyodon, Betula luminifera, Platycarya strobilacea; 15, Form. Quercus spinosa; 16, Form. Quercus glandulifera var. brevipetiolata; 17, Form. Castanea seguinii; 18, Form. Pinus henryi; 19, Form. Betula luminifera, Platycarya strobilacea, Carpinus sp.; 20, Form. Pinups armandii, Betula utilis; 21, Form. Quercus aliena var. acuteserrata; 22, Form. Abies chensiensis; 23, Form. Corylus heterophylla var. sutchuenensis, Viburnum opulus var. calvescens, Malus hupehensis; 24, Form. Carex sp., Sanguisorba officinalis var. longifolia, Anaphalis sinica, Geranium henryi; 25, Form. Rhus verniciflua, Quercus acutidentata, Cornus controversa, Platycarya strobilacea; 26, Form. Miscanthus sinensis, Pteridium aquilinum var. latiusculum; 27, Form. Lespedeza formosa, L. buergeri; 28, Form. Carex sp., Juncus concinus, Sanguisorba officicnalis var. longifolia, Chamaenerion angustifolium; 29, Form. Pinus armandii, Querus aliena var. acuteserrata; 30, Form. Pinus armandii; 31, Form. Fagus engleriana; 32, Form. Abies chensiensis, Picea wilsonii; 33, Form. Quercus aliena var. acuteserrata, Fagus engleriana, Betula albosinensis; 34, Form. Betula albosinensis; 35, Form. Pinus armandii, Populus davidiana, Betula albosinensis; 36, Form. Pinus armandii, Populus davidiana; 37, Form. Salix carprea var. inica, Crataegus wilsonii, Sorbus hupehensis; 38, Form. Abies fargesii, Betula albosinensis, Acer sp.; 39, Form. Salix cupularis; 40, Form. Berberis dasystachya; 41, Form. Sinarundinaria nitida; 42, Form. Cotoneaster horizontalis; 43, Form. Abies fargesii; 44, Form. Rhododendron fargesii; 45, Form. Sabina pingii var. wilsonii; 46, Form. Trisetum clarkei, Festuca rubra, Deyeuxia scabrescens. Formation distribution along the elevation was based on Tian (2002).
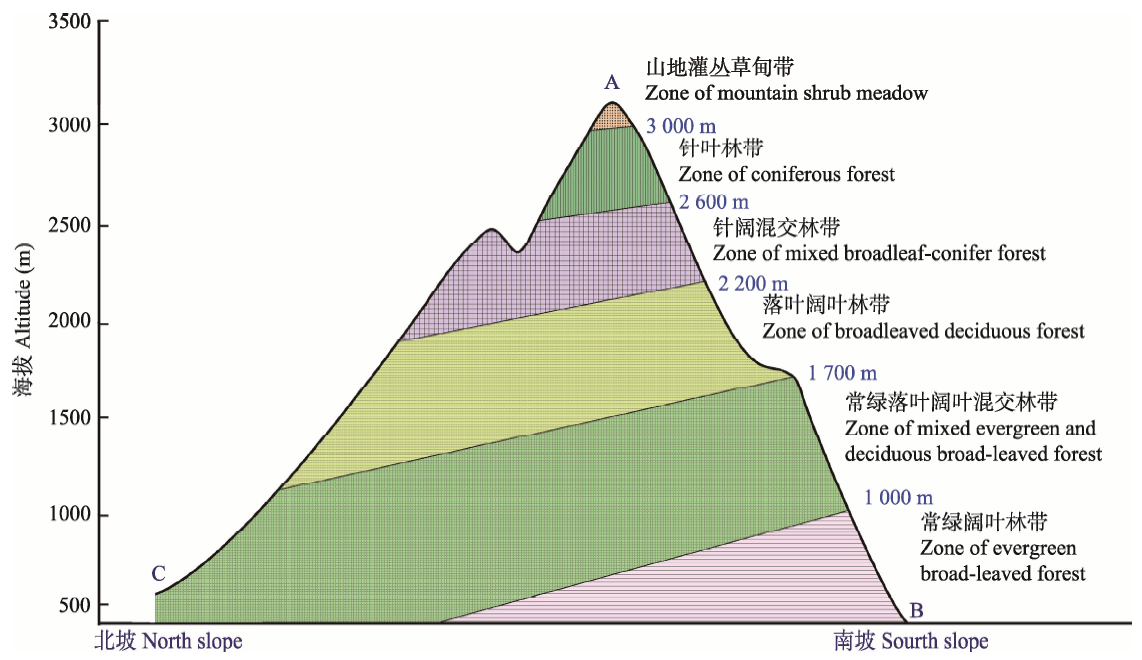
图3 神农架山地示意图。A, 神农顶, 神农架遗产地最高点, 海拔3 106.2 m; B, 下谷坪, 神农架遗产地南坡最低点, 海拔400 m; C, 韩家坪, 神农架遗产地北坡最低点, 海拔600 m。
Fig. 3 Sketch map of Shennongjia mountain. A, Shennongding, the summit of the heritage site with the elevation of 3 106.2 m; B, Xiaguping, the lowest point of the south slope of the heritage site with the elevation of 400 m; C, Hanjiaping, the lowest point of the north slope of the heritage site with the elevation of 600 m.
 |
表2 Udvardy东方落叶林生物地理省内主要山地的现存植被垂直带谱
Table 2 The altitudinal vegetation zonation of the Eastern Deciduous Forest Biogeographic Province in the Classification of the Biogeographical Provinces of the World by Udvardy
|
| [1] |
Bahram M, Polme S, Koljalg U, Zarre S, Tedersoo L (2012). Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytologist, 193, 465-473.
DOI URL PMID |
| [2] | Barthlott W, Lauer WA, Placke A (1996). Global distribution of species diversity in vascular Plants: Towards a world map of phytodiversity.Erdkunde, 50, 317-327. |
| [3] | Beniston M (2003). Climatic change in mountain regions: A review of possible impacts.Climatic Change, 59, 5-31.> |
| [4] |
Colwell RK, Brehm G, Cardelus CL, Gilman AC, Longino JT (2008). Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics.Science, 322, 258-261.
DOI URL PMID |
| [5] | Editorial Board of Anhui’s Forest (1990). Forest of Anhui. Anhui Scientific and Technical Press, Hefei. (in Chinese)[安徽森林编委会 (1990). 安徽森林. 安徽科学技术出版社, 合肥.] |
| [6] | Editorial Board of Anhui’s Vegetation (1983). Vegetation of Anhui. Anhui Scientific and Technical Press, Hefei. (in Chinese)[安徽植被编委会 (1983). 安徽植被. 安徽科学技术出版社, 合肥.] |
| [7] | Editorial Board of Guizhou’s Forest (1989). Forest of Guizhou, China Forestry Publishing House, Beijing. (in Chinese)[贵州森林编委会 (1989). 贵州森林. 中国林业出版社, 北京.] |
| [8] | Editorial Board of Hubei’s Forest (1990). Forest of Hubei. China Forestry Publishing House,, Wuhan. (in Chinese)[湖北森林编委会 (1990). 湖北森林. 中国林业出版社, 武汉.] |
| [9] | Editorial Board of Hunan’s Forest (1991). Forest of Hunan. Hunan Scientific and Technical Press, Changsha. (in Chinese)[湖南森林编委会 (1991). 湖南森林. 湖南科学技术出版社, 长沙.] |
| [10] | Editorial Board of Jiangxi’s Forest (1983). Forest of Jiangxi. China Forestry Publishing House, Beijing. (in Chinese)[江西森林编委会 (1983). 江西森林. 中国林业出版社, 北京.] |
| [11] | Board of Physical Geography of China, Chinese Academy of Sciences (1980). The Physical Geography of China (Volume of Physiognomy). Science Press, Beijing. (in Chinese)[中国科学院中国自然地理编委会 (1980). 中国自然地理·地貌卷. 科学出版社, 北京.] |
| [12] | Editorial Board of Shaanxi’s Forest (1989). Forest of Shaanxi. China Forestry Publishing House, Beijing. (in Chinese)[陕西森林编委会 (1989). 陕西森林. 中国林业出版社, 北京.] |
| [13] | Editorial Board of Sichuan’s Forest (1983). Forest of Sichuan, China Forestry Publishing House, Beijing. (in Chinese)[四川森林编委会 (1983). 四川森林. 中国林业出版社, 北京.] |
| [14] | Editorial Committee of Vegetation Map of China, Chinese Academy of Sciences (2007). Vegetation Map of People’s Republic of China. Geological Publishing House, Beijing. (in Chinese)[中国科学院中国植被图编辑委员会 (2007). 中国植被及其地理格局. 地质出版社, 北京.] |
| [15] |
Gomez-Hernandez M, Williams-Linera G, Guevara R, Lodge DJ (2012). Patterns of macromycete community assemblage along an elevation gradient: Options for fungal gradient and metacommunity analyses.Biodiversity and Conservation, 21, 2247-2268.
DOI URL |
| [16] |
Gonzalez LG, Geeraerad AH, Spilimbergo S, Elstk K, Ginneken LV, Debevere J, van Impe JF, Devlieghere F (2007). High pressure carbon dioxide inactivation of microorganisms in foods: The past, the present and the future.International Journal of Food Microbiology, 117, 1-28.
DOI URL PMID |
| [17] | Grinnell J (1924). Geography and evolution.Ecology, 5, 225-229. |
| [18] | He F (2006). Forest vegetation in Japan.Journal of Sichuan Forestry Science and Technology, (3), 38-41. (in Chinese with English abstract)[何飞 (2006). 日本的森林植被. 四川林业科技, (3), 38-41.] |
| [19] |
He X, Hou E, Wen D (2016). Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China.Scientific Reports, 6, 24261. doi: 10.1038/srep24261.
DOI URL PMID |
| [20] | Hodkinson ID (2005). Terrestrial insects along elevation gradients: Species and community responses to altitude.Biology Review, 80, 489-513. |
| [21] | Jiang MQ, Chen RJ, Sun YF (1982). The vegetation in Huangshan. Nature Journal, (3), 222-226, 197. (in Chinese with English abstract)[蒋木青, 陈仁钧, 孙毓飞 (1982). 黄山的植被. 自然杂志,(3), 222-226, 197.] |
| [22] | Korner C (2007). The use of “altitude” in ecological research.Trends in Ecology and Evolution, 22, 569-574. |
| [23] | Lei MD (1999). Vegetation of Shaanxi. Science Press, Beijing. (in Chinese)[雷明德 (1999). 陕西植被. 科学出版社, 北京.] |
| [24] | Li XD (1985). Research about vertical zonation of vegetation on the southern slope of the west Qinling.Shanxi Forest Science and Technology, (3), 88-92. (in Chinese with English abstract)[李晓东 (1985). 对陕西秦岭西段南坡植被垂直带划分问题的一点认识. 陕西林业科技, (3), 88-92.] |
| [25] |
Li YM, Xu L, Ma Y, Yang JY, Yang YH (2003). The species richness of nonvolant mammals in Shennongjia Nature Reserve, Hubei Province, China: Distribution patterns along elevational gradient. Biodiversity Science, 11, 1-9. (in Chinese with English abstract)[李义明, 许龙, 马勇, 杨敬元, 杨玉慧 (2003). 神农架自然保护区非飞行哺乳动物的物种丰富度: 沿海拔梯度的分布格局. 生物多样性, 11, 1-9.
DOI URL |
| [26] | Liao MY (2015). Comprehensive Survey Report of Natural Resources in Shennongjia Area. China Forestry Publishing House, Beijing. (in Chinese)[廖明尧 (2015). 神农架地区自然资源综合调查报告. 中国林业出版社, 北京.] |
| [27] |
Ma KP (2016). Conservation of world natural heritage should be ensured and its contribution to local sustainable development promoted.Biodiversity Science, 24, 861-862. (in Chinese with English abstract)[马克平 (2016). 世界自然遗产既要加强保护也要适度利用. 生物多样性, 24, 861-862.]
DOI URL |
| [28] |
Malhi Y, Silman M, Salinas N, Bush M, Meir P, Saatchi S (2010). Introduction: Elevation gradients in the tropics: Laboratories for ecosystem ecology and global change research.Global Change Biology, 16, 3171-3175.
DOI URL |
| [29] |
McCain CM (2009). Global analysis of bird elevational diversity.Global Ecology and Biogeography, 18, 346-360.
DOI URL |
| [30] |
Ohsawa M (1984). Differentiation of vegetation zones and species strategies in the subalpine region of Mt. Fuji.Vegetatio, 57, 15-52.
DOI URL |
| [31] | Olson DM (1994). The distribution of leaf litter invertebrates along a Neotropical altitudinal gradient.Journal of Tropical Ecology, 10, 129-150. |
| [32] |
Palin OF, Eggleton P, Malhi Y, Girardin CAJ, Rozas-D’avila A, Parr CL (2011). Termite diversity along an Amazon- Andes elevation gradient, Peru.Biotropica, 43, 100-107.
DOI URL |
| [33] | Qi CJ (1990). Hunan Vegetation. Hunan Scientific and Technical Press, Changsha. (in Chinese)[祁承经 (1990). 湖南植被. 湖南科学技术出版社, 长沙.] |
| [34] | Rahbek C (2005). The role of spatial scale and the perception of large-scale species-richness patterns.Ecology Letters, 8, 224-239. |
| [35] |
Raich JW, Russell AE, Vitousek PM (1997). Primary productivity and ecosystem development along an elevational gradient on Mauna Loa, Hawaii.Ecology, 78, 707-721.
DOI URL |
| [36] |
Salinas N, Malhi Y, Meir P, Silman M, Roman Cuesta R, Huaman J, Salinas D, Huaman V, Gibaja A, Mamani M, Farfan F (2011). The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests.New Phytologist, 189, 967-977.
DOI URL PMID |
| [37] |
Sanders NJ, Lessard JP, Fitzpatrick MC, Dunn RR (2007). Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains. Global Ecology and Biogeography, 16, 640-649.
DOI URL |
| [38] |
Sundqvist MK, Sanders NJ, Wardle DA (2013). Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change.Annual Review of Ecology, Evolution and Systematics, 44, 261-280.
DOI URL |
| [39] | Tan JS, Ban JD, Wang ZX (1982). The vegetation regionalization of Huibei Province.Journal of Central China Normal University (Natural Sciences), (3), 102-127. (in Chinese with English abstract)[谭景燊, 班继德, 王增学 (1982). 湖北植被区划. 华中师院学报(自然科学版), (3), 102-127.] |
| [40] | Tanner EVJ, Vitousek PM, Cuevas E (1998). Experimental investigation of nutrient limitation of forest growth on wet tropical mountains.Ecology, 79, 10-22. |
| [41] | Tian ZQ (2002). The Vegetation of Shennongjia and 1:200, 000 Vegetation Map. PhD dissertation, Institute of Botany, Chinese Academy of Sciences, Beijing. (in Chinese with English abstract)[田自强 (2002). 神农架的植被及其1:200, 000植被图的编制. 博士学位论文, 中国科学院植物研究所, 北京.] |
| [42] | Udvardy MDF (1975). A Classification of the Biogeographical Provinces of the World. IUCN Occasional Paper. . |
| [43] |
Vitousek PM, Matson PA, Turner DR (1988). Elevational and age gradients in Hawaiian montane rainforest: Foliar and soil nutrients.Oecologia, 77, 565-570.
DOI URL |
| [44] |
Wagai R, Kitayama K, Satomura T, Fujinuma R, Balser T (2011). Interactive influences of climate and parent material on soil microbial community structure in Bornean tropical forest ecosystems.Ecological Research, 26, 627-636.
DOI URL |
| [45] | Walter H (1984). World Vegetation. Translated by Institute of Botany, Chinese Academy of Sciences. Science Press, Beijing. (in Chinese)[沃尔特 (1984). 世界植被. 中国科学院植物研究所, 译. 科学出版社, 北京.] |
| [46] | Wang XP, Wang ZH, Fang JY (2004). Mountain ranges and peaks in China.Biodiversity Science, 12, 206-212. (in Chinese with English abstract)[王襄平, 王志恒, 方精云 (2004). 中国的主要山脉和山峰. 生物多样性, 12, 206-212.] |
| [47] | Wardle DA (2002). Communities and Ecosystems: Linking the Aboveground and Belowground Components. Princeton University Press, Princeton, USA. |
| [48] | Whittaker RH (1956). Vegetation of the Great Smokey Mountains.Ecological Monographys, 26, 1-80. |
| [49] | Whittaker RH (1960). Vegetation of the Siskiyou Mountains, Oregon and California.Ecological Monographys, 30, 279-338. |
| [50] | Wu GF, Zhou XJ, Tang YF (1988). The vegetation types and their distributions in Sanqingshan Jiangxi Province.Journal of East China Normal University (Natural Science), (2), 87-96. (in Chinese with English abstract)[吴国芳, 周秀佳, 汤艺峰 (1988). 江西省三清山的植被类型及其分布. 华东师范大学学报(自然科学版), (2), 87-96.] |
| [51] | Ying JS, Chen ML (2011). Plant Geography of China. Shanghai Scientific and Technical Publishers, Shanghai. (in Chinese)[应俊生, 陈梦玲 (2011). 中国植物地理. 上海科学技术出版社, 上海.] |
| [52] |
Zhang B (2014). Diversity of Bird Communities in Shennongjia National Nature Reserve. PhD dissertation, Central China Normal University, Wuhan. (in Chinese with English abstract)[章波 (2014). 神农架国家级自然保护区鸟类群落多样性研究. 博士学位论文, 华中师范大学, 武汉.]
DOI URL |
| [53] | Zhu ZQ, Song CS(1999). Scientific Survey of Shennongjia Nature Reserve. China Forestry Publishing House, Beijing. (in Chinese)[朱兆泉, 宋朝枢 (1999). 神农架自然保护区科学考察集. 中国林业出版社, 北京.] |
| [1] | 张尧, 陈岚, 王洁莹, 李益, 王俊, 郭垚鑫, 任成杰, 白红英, 孙昊田, 赵发珠. 太白山不同海拔森林根际土壤微生物碳利用效率差异性及其影响因素[J]. 植物生态学报, 2023, 47(2): 275-288. |
| [2] | 彭鑫, 金光泽. 植物特性和环境因子对阔叶红松林暗多样性的影响[J]. 植物生态学报, 2022, 46(6): 656-666. |
| [3] | 卢晶, 马宗祺, 高鹏斐, 樊宝丽, 孙坤. 祁连山区演替先锋物种西藏沙棘的种群结构及动态对海拔梯度的响应[J]. 植物生态学报, 2022, 46(5): 569-579. |
| [4] | 牟文博, 徐当会, 王谢军, 敬文茂, 张瑞英, 顾玉玲, 姚广前, 祁世华, 张龙, 苟亚飞. 排露沟流域不同海拔灌丛土壤碳氮磷化学计量特征[J]. 植物生态学报, 2022, 46(11): 1422-1431. |
| [5] | 解梦怡, 冯秀秀, 马寰菲, 胡汗, 王洁莹, 郭垚鑫, 任成杰, 王俊, 赵发珠. 秦岭锐齿栎林土壤酶活性与化学计量比变化特征及其影响因素[J]. 植物生态学报, 2020, 44(8): 885-894. |
| [6] | 王进, 朱江, 艾训儒, 姚兰, 黄小, 吴漫玲, 朱强, 洪建峰. 湖北星斗山地形变化对不同生活型植物叶功能性状的影响[J]. 植物生态学报, 2019, 43(5): 447-457. |
| [7] | 高思涵, 葛珏希, 周李奕, 朱宝琳, 葛星宇, 李凯, 倪健. 测定森林树木叶面积的最适叶片数是多少?[J]. 植物生态学报, 2018, 42(9): 917-925. |
| [8] | 周天阳, NARAYAN Prasad Gaire, 廖礼彬, 郑莉莉, 王金牛, 孙建, 魏彦强, 谢雨, 吴彦. 青藏高原东缘两处高山树线交错带时空动态及其建群种的生态学特征[J]. 植物生态学报, 2018, 42(11): 1082-1093. |
| [9] | 杨蕾, 孙晗, 樊艳文, 韩威, 曾令兵, 刘超, 王襄平. 长白山木本植物叶片氮磷含量的海拔梯度格局及影响因子[J]. 植物生态学报, 2017, 41(12): 1228-1238. |
| [10] | 于健, 徐倩倩, 刘文慧, 罗春旺, 杨君珑, 李俊清, 刘琪璟. 长白山东坡不同海拔长白落叶松径向生长对气候变化的响应[J]. 植物生态学报, 2016, 40(1): 24-35. |
| [11] | 黄永涛, 姚兰, 艾训儒, 吕世安, 丁易. 鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析[J]. 植物生态学报, 2015, 39(10): 990-1002. |
| [12] | 吴则焰,林文雄,陈志芳,方长旬,张志兴,吴林坤,周明明,陈婷. 中亚热带森林土壤微生物群落多样性随海拔梯度的变化[J]. 植物生态学报, 2013, 37(5): 397-406. |
| [13] | 徐波, 王金牛, 石福孙, 高景, 吴宁. 青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应[J]. 植物生态学报, 2013, 37(3): 187-196. |
| [14] | 张文涛, 江源, 王明昌, 张凌楠, 董满宇, 郭媛媛. 芦芽山阳坡不同海拔白杄径向生长对气候变暖的响应[J]. 植物生态学报, 2013, 37(12): 1142-1152. |
| [15] | 彭剑峰, 勾晓华, 陈发虎, 方克艳, 张芬. 坡向对海拔梯度上祁连圆柏树木生长的影响[J]. 植物生态学报, 2010, 34(5): 517-525. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19