
植物生态学报 ›› 2018, Vol. 42 ›› Issue (1): 38-49.DOI: 10.17521/cjpe.2017.0255
所属专题: 青藏高原植物生态学:群落生态学
字洪标,陈焱,胡雷,王长庭( )
)
出版日期:2018-01-20
发布日期:2018-03-08
通讯作者:
王长庭
基金资助:
ZI Hong-Biao,CHEN Yan,HU Lei,WANG Chang-Ting()
Online:2018-01-20
Published:2018-03-08
Contact:
Chang-Ting WANG
Supported by:摘要:
该文以川西北高寒草甸为研究对象, 采用微根管法研究了不同施氮(N)水平下高寒草甸植物群落根系现存量、生产量、死亡量和周转率的变化及其与土壤理化性质的相互关系。结果表明: N添加显著增加了土壤速效氮(AN)含量, 降低了土壤pH值, 但是对土壤有机质(SOM)和全氮(TN)含量无显著影响。在0-10 cm土层, 平均根系现存量和累积根系生产量无显著变化, 累积根系死亡量在N10处理下显著降低了206.1 g·m -2, 根系周转率在N30处理下显著提高了17%; 在10-20 cm土层, N添加处理的平均根系现存量和累积根系生产量分别显著降低了195.3和142.3 g·m -2 (N10)、235.8和212.1 g·m -2 (N20)、198.0和204.4 g·m -2 (N30), 累积根系死亡量和周转率无显著变化。此外, 累积根系生产量、死亡量和周转率与AN含量相关性较大, 而平均根系现存量与SOM、AN和TN含量相关性较大。综上所述, N添加对高寒草甸的影响主要通过改变土壤可利用N含量, 进而影响根系的动态特征、空间分布格局和周转以及碳分配特征。
字洪标, 陈焱, 胡雷, 王长庭. 氮肥添加对川西北高寒草甸植物群落根系动态的影响. 植物生态学报, 2018, 42(1): 38-49. DOI: 10.17521/cjpe.2017.0255
ZI Hong-Biao, CHEN Yan, HU Lei, WANG Chang-Ting. Effects of nitrogen addition on root dynamics in an alpine meadow, Northwestern Sichuan. Chinese Journal of Plant Ecology, 2018, 42(1): 38-49. DOI: 10.17521/cjpe.2017.0255
| 土层 Soil layer (cm) | 处理 Treatment | pH值 pH value | 有机质 SOM (g·kg-1) | 速效氮 AN (mg·kg-1) | 全氮 TN (g·kg-1) |
|---|---|---|---|---|---|
| 0-10 cm | CK | 5.74 ± 0.10Aa | 110.92 ± 10.48Aa | 343.00 ± 9.40Ac | 6.38 ± 0.84Aa |
| N10 | 5.42 ± 0.14Bb | 119.61 ± 19.21Aa | 323.40 ± 9.96Ad | 7.01 ± 0.81Aa | |
| N20 | 5.50 ± 0.25Ab | 121.68 ± 25.26Aa | 392.00 ± 2.46Ab | 6.95 ± 0.70Aa | |
| N30 | 5.44 ± 0.08Ab | 124.58 ± 11.78Aa | 418.95 ± 9.15Aa | 7.27 ± 0.97Aa | |
| 10-20 cm | CK | 5.73 ± 0.08Aa | 94.60 ± 11.23Aa | 290.33 ± 7.99Bb | 5.34 ± 0.93Aa |
| N10 | 5.69 ± 0.27Aa | 88.10 ± 24.43Ba | 276.85 ± 8.20Bb | 5.20 ± 0.71Aa | |
| N20 | 5.62 ± 0.19Ab | 91.91 ± 17.75Ba | 328.30 ± 6.12Ba | 5.40 ± 0.83Aa | |
| N30 | 5.48 ± 0.16Ab | 95.30 ± 16.64Ba | 316.05 ± 4.85Ba | 5.64 ± 0.85Aa |
表1 氮添加梯度对寒草甸土壤养分的影响(平均值±标准偏差, n = 6)
Table 1 Effect of nitrogen addition on soil nutrients of alpine meadow (mean ± SD, n = 6)
| 土层 Soil layer (cm) | 处理 Treatment | pH值 pH value | 有机质 SOM (g·kg-1) | 速效氮 AN (mg·kg-1) | 全氮 TN (g·kg-1) |
|---|---|---|---|---|---|
| 0-10 cm | CK | 5.74 ± 0.10Aa | 110.92 ± 10.48Aa | 343.00 ± 9.40Ac | 6.38 ± 0.84Aa |
| N10 | 5.42 ± 0.14Bb | 119.61 ± 19.21Aa | 323.40 ± 9.96Ad | 7.01 ± 0.81Aa | |
| N20 | 5.50 ± 0.25Ab | 121.68 ± 25.26Aa | 392.00 ± 2.46Ab | 6.95 ± 0.70Aa | |
| N30 | 5.44 ± 0.08Ab | 124.58 ± 11.78Aa | 418.95 ± 9.15Aa | 7.27 ± 0.97Aa | |
| 10-20 cm | CK | 5.73 ± 0.08Aa | 94.60 ± 11.23Aa | 290.33 ± 7.99Bb | 5.34 ± 0.93Aa |
| N10 | 5.69 ± 0.27Aa | 88.10 ± 24.43Ba | 276.85 ± 8.20Bb | 5.20 ± 0.71Aa | |
| N20 | 5.62 ± 0.19Ab | 91.91 ± 17.75Ba | 328.30 ± 6.12Ba | 5.40 ± 0.83Aa | |
| N30 | 5.48 ± 0.16Ab | 95.30 ± 16.64Ba | 316.05 ± 4.85Ba | 5.64 ± 0.85Aa |
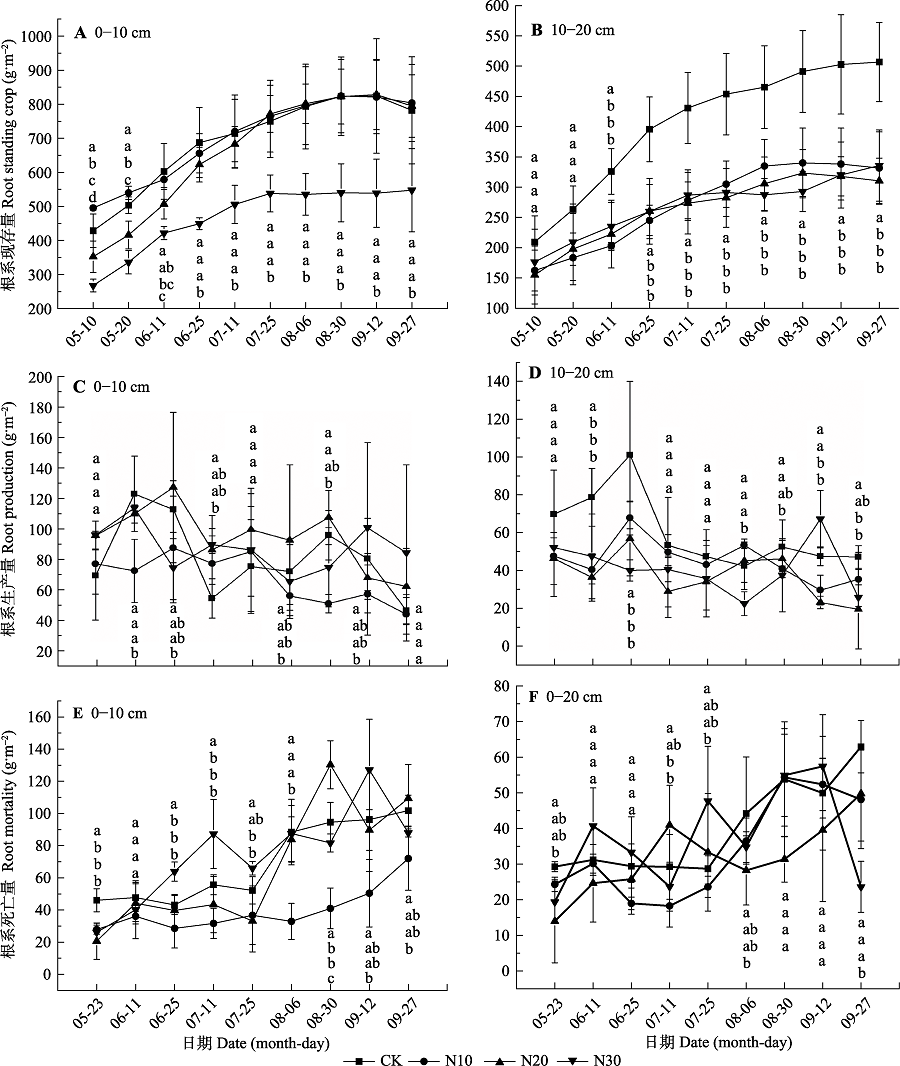
图1 不同氮添加梯度根系的季节动态(平均值±标准偏差, n = 6)。不同小写字母表示不同处理间差异显著(p < 0.05)。CK、N10、N20、N30同表1.
Fig. 1 Seasonal patterns of root for different nitrogen addition gradients (mean ± SD, n = 6). Different lowercase letters indicate significant differences in different treatment (p < 0.05). CK, N10, N20, N30, see Table 1.
| 土层 Soil layer | 根系指标 Root index | 取样时间 Sampling time | N添加梯度 N addition gradient | 取样时间×N添加梯度 Sampling time × N addition gradient | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| 0-10 cm | 根系现存量 Root standing crop | 81.262 | < 0.001 | 7.468 | 0.010 | 1.579 | 0.239 |
| 根系生产量 Root production | 63.809 | < 0.001 | 1.078 | 0.412 | 9.53 | < 0.001 | |
| 根系死亡量 Root mortality | 131.752 | < 0.001 | 4.477 | 0.04 | 9.693 | 0.001 | |
| 10-20 cm | 根系现存量 Root standing crop | 104.472 | < 0.001 | 7.793 | 0.009 | 3.918 | 0.250 |
| 根系生产量 Root production | 60.200 | < 0.001 | 3.460 | 0.071 | 12.932 | < 0.001 | |
| 根系死亡量 Root mortality | 138.540 | < 0.001 | 0.482 | 0.704 | 14.224 | < 0.001 | |
表2 不同N添加梯度根系动态的重复测量方差分析(n = 6)
Table 2 Repeated-measure variance analysis on the dynamic of plant roots of the different N addition fertilizer gradients (n = 6)
| 土层 Soil layer | 根系指标 Root index | 取样时间 Sampling time | N添加梯度 N addition gradient | 取样时间×N添加梯度 Sampling time × N addition gradient | |||
|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | ||
| 0-10 cm | 根系现存量 Root standing crop | 81.262 | < 0.001 | 7.468 | 0.010 | 1.579 | 0.239 |
| 根系生产量 Root production | 63.809 | < 0.001 | 1.078 | 0.412 | 9.53 | < 0.001 | |
| 根系死亡量 Root mortality | 131.752 | < 0.001 | 4.477 | 0.04 | 9.693 | 0.001 | |
| 10-20 cm | 根系现存量 Root standing crop | 104.472 | < 0.001 | 7.793 | 0.009 | 3.918 | 0.250 |
| 根系生产量 Root production | 60.200 | < 0.001 | 3.460 | 0.071 | 12.932 | < 0.001 | |
| 根系死亡量 Root mortality | 138.540 | < 0.001 | 0.482 | 0.704 | 14.224 | < 0.001 | |
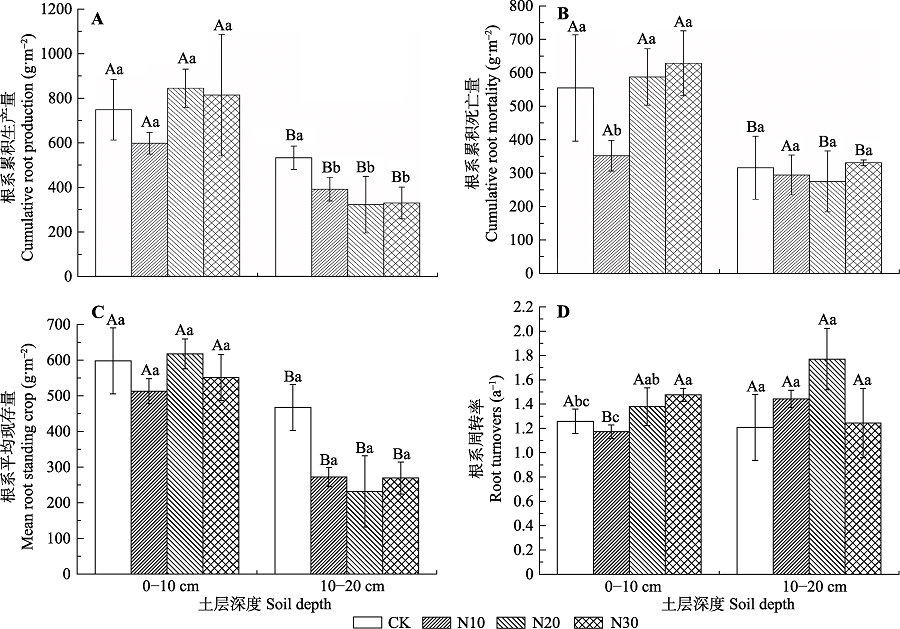
图2 不同氮添加梯度根系年生产量和死亡量、平均现存量和周转率的变化(平均值±标准差, n = 6)。不同小写字母表示不同处理间差异显著(p < 0.05); 不同大写字母表示不同土层间差异显著(p < 0.05)。CK、N10、N20、N30同表1。
Fig. 2 Cumulative length production, cumulative length mortality, mean root standing crop and Annual root turnovers over the growing season different nitrogen addition gradients (mean ± SD, n = 6). Different lowercase letters indicate significant differences in different treatment (p < 0.05). Different capital letters indicate significant different in different soil layers (p < 0.05). CK, N10, N20, N30, see Table 1.
| 因子 Factor | 处理 Treatment | 深度 Depth | 处理×深度 Treatment × Depth | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| 累积根系生产量 Cumulative root production | 1.328 | 0.300 | 47.14 | < 0.001 | 2.661 | 0.083 |
| 平均根系现存量 Mean root standing crop | 1.770 | 0.190 | 29.93 | < 0.001 | 1.237 | 0.329 |
| 根系周转 Root turnovers | 1.655 | 0.220 | 0.689 | 0.420 | 1.573 | 0.235 |
| 累积根系死亡量 Cumulative root mortality | 3.280 | 0.050 | 38.11 | < 0.001 | 2.538 | 0.093 |
表3 不同N添加梯度和土层深度对根系特征的方差分析(n = 6)
Table 3 ANOVA of on the dynamic of plant roots of the different N addition gradients (n = 6)
| 因子 Factor | 处理 Treatment | 深度 Depth | 处理×深度 Treatment × Depth | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| 累积根系生产量 Cumulative root production | 1.328 | 0.300 | 47.14 | < 0.001 | 2.661 | 0.083 |
| 平均根系现存量 Mean root standing crop | 1.770 | 0.190 | 29.93 | < 0.001 | 1.237 | 0.329 |
| 根系周转 Root turnovers | 1.655 | 0.220 | 0.689 | 0.420 | 1.573 | 0.235 |
| 累积根系死亡量 Cumulative root mortality | 3.280 | 0.050 | 38.11 | < 0.001 | 2.538 | 0.093 |
| 土层 Soil layer | 项目名称 Item | pH值 pH value | 有机质 SOM (g·kg-1) | 速效氮 AN (mg·kg-1) | 全氮 TN (mg·kg-1) |
|---|---|---|---|---|---|
| 0-10 cm | 根系现存量 Root standing crop | -0.27 | -0.414 | 0.487 | 0.622* |
| 根系生产量 Root production | 0.280 | -0.421 | -0.843** | -0.167 | |
| 根系死亡量 Root mortality | 0.315 | -0.044 | 0.716** | 0.021 | |
| 根系周转率 Root turnover | -0.459 | -0.225 | 0.673* | 0.691* | |
| 10-20 cm | 根系现存量 Root standing crop | -0.075 | -0.590* | -0.684* | 0.624* |
| 根系生产量 Root production | 0.371 | 0.005 | -0.413 | 0.095 | |
| 根系死亡量 Root mortality | -0.095 | -0.038 | -0.404 | 0.300 | |
| 根系周转率 Root turnover | -0.193 | -0.476 | 0.287 | 0.293 |
表4 土壤理化性质与根系特征的相关性(n = 6)
Table 4 Pearson correlation analysis between soil physicochemical properties and root characteristics (n = 6)
| 土层 Soil layer | 项目名称 Item | pH值 pH value | 有机质 SOM (g·kg-1) | 速效氮 AN (mg·kg-1) | 全氮 TN (mg·kg-1) |
|---|---|---|---|---|---|
| 0-10 cm | 根系现存量 Root standing crop | -0.27 | -0.414 | 0.487 | 0.622* |
| 根系生产量 Root production | 0.280 | -0.421 | -0.843** | -0.167 | |
| 根系死亡量 Root mortality | 0.315 | -0.044 | 0.716** | 0.021 | |
| 根系周转率 Root turnover | -0.459 | -0.225 | 0.673* | 0.691* | |
| 10-20 cm | 根系现存量 Root standing crop | -0.075 | -0.590* | -0.684* | 0.624* |
| 根系生产量 Root production | 0.371 | 0.005 | -0.413 | 0.095 | |
| 根系死亡量 Root mortality | -0.095 | -0.038 | -0.404 | 0.300 | |
| 根系周转率 Root turnover | -0.193 | -0.476 | 0.287 | 0.293 |
| [1] |
Anderson LJ, Comas LH, Lakso AN, Eissenstat DM ( 2003). Multiple risk factors in root survivorship: A 4-year study in Concord grape. New Phytologist, 158, 489-501.
DOI URL |
| [2] | Bai WM, Wan SQ, Niu SL, Liu WX, Chen QS, Wang QB, Zhang WH, Han XG, Li LH ( 2010). Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling. Global Change Biology, 6, 1306-1316. |
| [3] |
Bai WM, Wang ZW, Chen QS, Zhang WH, Li LH ( 2008). Spatial and temporal effects of nitrogen addition on root life span of Leymus chinensis in a typical steppe of Inner Mongolia. Functional Ecology, 22, 583-591.
DOI URL |
| [4] |
Bessler H, Temperton VM, Roscher C, Buchmann N, Schmid B, Schulze ED, Weisser WW, Engels C ( 2009). Aboveground overyielding in grassland mixtures is associated with reduced biomass partitioning to belowground organs. Ecology, 90, 1520-1530.
DOI URL PMID |
| [5] |
Bin ZJ, Wang JJ, Zhang WP, Xu DH, Cheng XH, Li KJ, Cao DH ( 2014). Effects of N addition on ecological stoichiometric characteristics in six dominant plant species of alpine meadow on the Qinghai-Xizang Plateau, China. Chinese Journal of Plant Ecology, 38, 231-237.
DOI URL |
|
[ 宾振钧, 王静静, 张文鹏, 徐当会, 程雪寒, 李柯杰, 曹德昊 ( 2014). 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响. 植物生态学报, 38, 231-237.]
DOI URL |
|
| [6] |
Birkhofer K, Bezemer TM, Bloem J, Bonkowski M, Christensen S, Dubois D, Ekelund F, Flie?bach A, Gunst L, Hedlund K, M?der P, Mikola J, Robin C, Set?l? H, Tatin-Froux F, van der Putten WH, Scheu S ( 2008). Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity. Soil Biology and Biochemistry, 40, 2297-2308.
DOI URL |
| [7] |
Cammeraat ELH, Risch AC ( 2008). The impact of ants on mineral soil properties and processes at different spatial scales. Journal of Applied Entomology, 132, 285-294.
DOI URL |
| [8] |
Chen D, Bai Y ( 2013). Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe. Journal of Ecology, 101, 1322-1334.
DOI URL |
| [9] |
Eissenstat DM, Wells CE, Yanai RD ( 2000). Building roots in a changing environment: Implications for root longevity. New Phytologist, 147, 33-42.
DOI URL |
| [10] |
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP, Asner GP, Cleveland CC, Green PA, Holland EA ( 2004). Nitrogen cycles: Past, present, and future. Biogeochemistry, 70, 153-226.
DOI URL |
| [11] |
Gaudinski JB, Trumbore SE, Davidson EA, Cook AC, Markwewitz D, Richter DD ( 2001). The age of fine root carbon in three forests of the eastern United States measured by radiocarbon. Oecologia, 129, 420-429.
DOI URL |
| [12] |
Gill RA, Jackson RB ( 2000). Global patterns of root turnover for terrestrial ecosystems. New Phytologist, 147, 13-31.
DOI URL |
| [13] | Guo DL, Fan PP ( 2007). Four hypotheses about the effects of soil nitrogen availability on fine root production and turnover. Chinese Journal of Applied Ecology, 18, 2354-2360. |
| [ 郭大立, 范萍萍 ( 2007). 关于氮有效性影响细根生产量和周转率的四个假说. 应用生态学报, 18, 2354-2360.] | |
| [14] |
Han WX, Fang JY, Guo DL, Zhang Y ( 2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 168, 377-385.
DOI URL PMID |
| [15] |
Hendricks JJ, Hendrick RL, Wilson CA, Mitchell RJ, Pecot SD, Guo D ( 2006). Assessing the patterns and controls of fine root dynamics: An empirical test and methodological review. Journal of Ecology, 94, 40-57.
DOI URL |
| [16] | Institute of Soil Science, Chinese Academy of Sciences ( 1983). Analysis of Soil Physical-Chemical Feature. Shanghai Science and Technology Press, Shanghai. |
| [ 中国科学院南京土壤研究所 ( 1983). 土壤理化分析. 上海科学技术出版社, 上海.] | |
| [17] |
Jackson RB, Mooney HA, Schulze ED ( 1997). A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences of the United States of America, 94, 7362-7366.
DOI URL PMID |
| [18] | Kong BB, Wei XH, Du JL, Li YN, Zhu ZH ( 2016). Effects of clipping and fertilization on the temporal dynamics of species diversity and functional diversity and their relationships in an alpine meadow. Chinese Journal of Plant Ecology, 40, 187-199. |
| [ 孔彬彬, 卫欣华, 杜家丽, 李英年, 朱志红 ( 2016). 刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响. 植物生态学报, 40, 187-199.] | |
| [19] |
Kurz I, O’Reilly CD, Tunney H ( 2006). Impact of cattle on soil physical properties and nutrient concentrations in overland flow from pasture in Ireland. Agriculture Ecosystems and Environment, 113, 378-390.
DOI URL |
| [20] |
Li WB, Jin CJ, Guan DX, Wang QK, Wang AZ, Yuan FH, Wu JB ( 2015). The effects of simulated nitrogen deposition on plant root traits: A meta-analysis. Soil Biology & Biochemistry, 82, 112-118.
DOI URL |
| [21] | Linderman RG ( 1994). Role of VAM fungi in biocontrol. In: Fleger FLP, Linderman RG eds. Mycorrhizae and Plant Health. American Phytopathological Society, St. Paul, USA. 1-25. |
| [22] |
Luo P, Han X, Wang Y, Han M, Shi H, Liu N, Bai HZ ( 2015) Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China. Annals of Microbiology, 65, 533-542.
DOI URL PMID |
| [23] |
Macdonald JA, Dise NB, Matzner E, Armbruster M, Gundersen P, Forsius M ( 2002). Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests. Global Change Biology, 8, 1028-1033.
DOI URL |
| [24] | Majdi H, Andersson P ( 2005). Fine root production and turnover in a Norway spruce stand in northern Sweden: Effects of nitrogen and water manipulation. Ecosystems, 8, 191-199. |
| [25] |
Majdi H, Ohrvik J ( 2004). Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden. Global Change Biology, 10, 182-188.
DOI URL |
| [26] |
Martin D, Chambers J ( 2002). Restoration of riparian meadows degraded by livestock grazing: Above- and belowground responses. Plant Ecology, 163, 77-91.
DOI URL |
| [27] |
Mei L, Wang ZQ, Cheng YH, Guo DL ( 2004). A review: Factors influencing fine root longevity in forest ecosystems. Acta Phytoecologica Sinica, 28, 704-710.
DOI URL |
|
[ 梅莉, 王政权, 程云环, 郭大立 ( 2004). 林木细根寿命及其影响因子研究进展. 植物生态学报, 28, 704-710.]
DOI URL |
|
| [28] | Mei L, Wang ZQ, Zhang XJ, Yu LZ, Du Y ( 2008). Effects of nitrogen fertilization on fine root biomass production and turnover of Fraxinus mandshurica plantation. Chinese Journal of Ecology, 27, 1663-1668. |
| [ 梅莉, 王政权, 张秀娟, 于立忠, 杜英 ( 2008). 施氮肥对水曲柳人工林细根生产和周转的影响. 生态学杂志, 27, 1663-1668.] | |
| [29] |
Müller I, Schmid B, Weiner J ( 2000). The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology Evolution and Systematics, 3, 115-127.
DOI URL |
| [30] |
Nadelhoffer KJ, Norby R, Fitter A, Jackson R ( 2000). The potential effects of nitrogen deposition on fine-root production in forest ecosystems. New Phytologist, 147, 131-139.
DOI URL |
| [31] |
Norby RJ, Joanne L, Reilly CD, Miller NE, O’Neill EG ( 2004). Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. Proceedings of the National Academy of Sciences of the United States of America, 101, 9689-9693.
DOI URL |
| [32] | Pei ZQ, Yong Z, Zheng YR, Xiao CW ( 2011). Contribution of fine root turnover to the soil organic carbon cycling in a Reaumuria soongorica community in an arid ecosystem of Xinjiang Uygur Autonomous Region, China. Chinese Journal of Plant Ecology, 35, 1182-1191. |
| [ 裴智琴, 周勇, 郑元润, 肖春旺 ( 2011). 干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献. 植物生态学报, 35, 1182-1191.] | |
| [33] |
Peng Q, Dong Y, Qi YC ( 2008). Influence of external nitrogen input on key processes of carbon cycle in terrestrial ecosystem. Advances in Earth Science, 23, 874-883.
DOI URL |
|
[ 彭琴, 董云社, 齐玉春 ( 2008). 氮输入对陆地生态系统碳循环关键过程的影响. 地球科学进展, 23, 874-883.]
DOI URL |
|
| [34] |
Pregitzer KS, Burton AJ, Zak DR, Talhelm AF ( 2008). Simulated chronic nitrogen deposition increases carbon storage in northern temperate forests. Global Change Biology, 14, 142-153.
DOI URL |
| [35] |
Pregitzer KS, Zak DR, Curtis PS, Kubiske ME, Teeri JA, Vogel CS ( 1995). Atmospheric CO2, soil nitrogen and turnover of fine roots. New Phytologist, 129, 579-585.
DOI URL |
| [36] |
Rasse DP ( 2002). Nitrogen deposition and atmospheric CO2 interactions on fine root dynamics in temperate forests: A theoretical model analysis. Global Change Biology, 8, 486-503.
DOI URL |
| [37] |
Reay DS, Dentener F, Smith P, Grace J, Feely RA ( 2008). Global nitrogen deposition and carbon sinks. Nature Geoscience, 1, 430-437.
DOI URL |
| [38] |
Ren ZW, Li Q, Chu CJ, Zhao LQ, Zhang JQ, Ai DXC, Yang YB, Wang G ( 2010). Effects of resource additions on species richness and ANPP in an alpine meadow community. Journal of Plant Ecology, 3, 25-31.
DOI URL |
| [39] |
Rytter RM, Rytter L ( 2012). Quantitative estimates of root densities at minirhizotrons differ from those in the bulk soil. Plant and Soil, 350, 205-220.
DOI URL |
| [40] |
Sanders JL, Brown DA ( 1978). A new fiber optic technique for measuring root growth of soybeans under field conditions. Agronomy Journal, 70, 1073-1076.
DOI URL |
| [41] | Schoettle AW, Fahey T ( 1994). Foliage and fine root longevity in pines. Ecological Bulletins, 43, 136-153. |
| [42] |
Song MH, Yu FH, Ouyang H, Cao GM, Xu XL, Cornerlissen JHC ( 2012). Different inter-annual responses to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing. Global Change Biology, 18, 3100-3111.
DOI URL PMID |
| [43] |
Taylor HM, Ruck MG, Klepper B, Lund ZF ( 1970). Measurement of soil-grown roots in a rhizotronl. Agronomy Journal, 62, 807-809.
DOI URL |
| [44] |
Thomas RQ, Canham CD, Weathers KC, Goodale CL ( 2010). Increased tree carbon storage in response to nitrogen deposition in the US. Nature Geoscience, 3, 13-17.
DOI URL |
| [45] |
Tierney GL, Fahey TJ ( 2001). Evaluating minirhizotron estimates of fine root longevity and production in the forest floor of a temperate broadleaf forest. Plant and Soil, 229, 167-176.
DOI URL |
| [46] | Tu LH, Chen G, Peng Y, Hu HL, Hu TX, Zhang J ( 2014). Response of fine root decomposition to simulated nitrogen deposition in Pleioblastus amarus plantation, rainy area of west China. Chinese Journal of Applied Ecology, 25, 2176-2182. |
| [ 涂利华, 陈刚, 彭勇, 胡红玲, 胡庭兴, 张健 ( 2014). 华西雨屏区苦竹细根分解对模拟氮沉降的响应. 应用生态学报, 25, 2176-2182.] | |
| [47] |
van der Krift TAJ, Berendse F ( 2002). Root life spans of four grass species from habitats differing in nutrient availability. Functional Ecology, 16, 198-203.
DOI URL |
| [48] |
Vogt KA, Grier CC, Vogt DJ ( 1986). Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests. Advances in Ecological Research, 15, 303-377.
DOI URL |
| [49] |
Vogt KA, Vogt DJ, Palmiotto PA, Boon P, O’Hara J, Asbjornsen H ( 1995). Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. Plant and Soil, 187, 159-219.
DOI URL |
| [50] |
Wan SQ, Norby RJ, Pregitzer KS, Ledford J, O’Neill EG ( 2004). CO2 enrichment and warming of the atmosphere enhance both productivity and mortality of maple tree fine roots. New Phytologist, 162, 437-446.
DOI URL |
| [51] |
Wang CT, Long RJ, Wang QL, Liu W, Jing ZC, Zhang L ( 2010). Fertilization and litter effects on the functional group biomass, species diversity of plants, microbial biomass, and enzyme activity of two alpine meadow communities. Plant and Soil, 331, 377-389.
DOI URL |
| [52] |
Wang CT, Wang GX, Liu W, Wang QL ( 2013). Effects of fertilization gradients on plant community structure and soil characteristics in alpine meadow. Acta Ecologica Sinica, 33, 3103-3113.
DOI URL |
|
[ 王长庭, 王根绪, 刘伟, 王启兰 ( 2013). 施肥梯度对高寒草甸群落结构、功能和土壤质量的影响. 生态学报, 33, 3103-3113.]
DOI URL |
|
| [53] |
Wang R, Balkanski Y, Boucher O, Ciais P, Pe?uelas J, Tao S ( 2015). Significant contribution of combustion-related emissions to the atmospheric phosphorus budget. Nature Geoscience, 8, 48-54.
DOI URL |
| [54] |
Wei C, Huang QT, Hu SR, Zhang HQ, Zhang WX, Wang ZM, Zhu MG, Dai PW, Huang LZ ( 2014). Simultaneous electrochemical determination of hydroquinone, catechol and resorcinol at Nafion/multi-walled carbon nanotubes/carbon dots/multi-walled carbon nanotubes modified glassy carbon electrode. Electrochimica Acta, 149, 237-244.
DOI URL |
| [55] |
Woodward FI, Osborne CP ( 2000). The representation of root processes in models addressing the responses of vegetation to global change. New Phytologist, 147, 223-232.
DOI URL |
| [56] |
Wu YB, Che RX, Ma S, Deng YC, Zhu MJ, Cui XY ( 2010). Estimation of root production and turnover in an alpine meadow: Comparison of three measurement methods. Acta Ecologica Sinica, 34, 3529-3537.
DOI URL |
|
[ 吴伊波, 车荣晓, 马双, 邓永翠, 朱敏健, 崔骁勇 ( 2014). 高寒草甸植被细根生产和周转的比较研究. 生态学报, 34, 3529-3537.]
DOI URL |
|
| [57] | Xiong SP, Wang XC, Li CM, Ma XM, Du SY, Zhang YW, Ling SZ ( 2011). Responses of the spatial-temporal distribution of winter wheat (Triticum aestivum) roots and yield to different ratios of nitrogen sources. Chinese Journal of Plant Ecology, 35, 759-768. |
| [ 熊淑萍, 王小纯, 李春明, 马新明, 杜少勇, 张营武, 蔺世召 ( 2011). 冬小麦根系时空分布动态及产量对不同氮源配施的响应. 植物生态学报, 35, 759-768.] | |
| [58] |
Yang XX, Ren F, Zhou HK, He JS ( 2014). Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology, 38, 159-166.
DOI URL |
|
[ 杨晓霞, 任飞, 周华坤, 贺金生 ( 2014). 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应. 植物生态学报, 38, 159-166.]
DOI URL |
|
| [59] |
Yang YH, Fang JY, Tang YH, Ji CJ, Zheng CY, He JS, Zhu B ( 2008). Storage, patterns and controls of soil organic carbon in the Tibetan grassland. Global Change Biology, 14, 1592-1599.
DOI URL |
| [60] |
Yang ZL, van Ruijven J, Du GZ ( 2011). The effects of long-term fertilization on the temporal stability of alpine meadow communities. Plant and Soil, 345, 315-324.
DOI URL |
| [61] | Yu SQ, Wang ZQ, Shi JW, Quan XK, Mei L, Sun Y, Jia SX, Yu LJ ( 2007). Estimating fine-root longevity of Fraxinus mandshurica and Larix gmelinii using minirhizotrons. Journal of Plant Ecology (Chinese Version), 31, 102-109. |
| [ 于水强, 王政权, 史建伟, 全先奎, 梅莉, 孙玥, 贾淑霞, 于立中 ( 2007). 水曲柳和落叶松细根寿命的估计. 植物生态学报, 31, 102-109.] | |
| [62] |
Zhang JQ, Li Q, Ren ZW, Yang X, Wang G ( 2010). Effects of nitrogen addition on species richness and relationship between species richness and aboveground productivity of alpine meadow of the Qinghai-Tibetan Plateau, China. Chinese Journal of Plant Ecology, 23, 33-38.
DOI URL |
|
[ 张杰琦, 李奇, 任正炜, 杨雪, 王刚 ( 2010). 氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响. 植物生态学报, 34, 1125-1131.]
DOI URL |
|
| [63] |
Zhao XC, Lai LM, Zhu LH, Wang JJ, Wang YJ, Zhou JH, Jiang LH, Ma YJ, Zhao CQ, Zhen YR ( 2014). Fine root biomass, decomposition and turnover of Reaumuria soongorica communities in the Sangong River Basin. Acta Ecologica Sinica, 34, 4295-4303.
DOI URL |
|
[ 赵学春, 来利明, 朱林海, 王健健, 王永吉, 周继华, 姜联合, 马远见, 赵春强, 郑元润 ( 2014). 三工河流域两种琵琶柴群落细根生物量、分解与周转. 生态学报, 34, 4295-4303.]
DOI URL |
|
| [64] | Zhou XM ( 2001). Chinese Kobresia Meadows. Science Press, Beijing. |
| [ 周兴民 ( 2001). 中国嵩草草甸. 科学出版社, 北京.] |
| [1] | 李红琴, 张法伟, 仪律北. 高寒草甸表层土壤和优势植物叶片的化学计量特征对降水改变和氮添加的响应[J]. 植物生态学报, 2023, 47(7): 922-931. |
| [2] | 吕自立, 刘彬, 常凤, 马紫荆, 曹秋梅. 巴音布鲁克高寒草甸植物功能多样性与生态系统多功能性关系沿海拔梯度的变化[J]. 植物生态学报, 2023, 47(6): 822-832. |
| [3] | 李伟, 张荣. 亚高寒草甸群落结构决定群落生产力实例验证[J]. 植物生态学报, 2023, 47(5): 713-723. |
| [4] | 刘艳方, 王文颖, 索南吉, 周华坤, 毛旭锋, 王世雄, 陈哲. 青海海北植物群落类型与土壤线虫群落相互关系[J]. 植物生态学报, 2022, 46(1): 27-39. |
| [5] | 董利军, 李金花, 陈珊, 张瑞, 孙建, 马妙君. 若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析[J]. 植物生态学报, 2021, 45(5): 507-515. |
| [6] | 汪子微, 万松泽, 蒋洪毛, 胡扬, 马书琴, 陈有超, 鲁旭阳. 青藏高原不同高寒草地类型土壤酶活性及其影响因子[J]. 植物生态学报, 2021, 45(5): 528-538. |
| [7] | 马书琴, 汪子微, 陈有超, 鲁旭阳. 藏北高寒草地土壤有机质化学组成对土壤蛋白酶和脲酶活性的影响[J]. 植物生态学报, 2021, 45(5): 516-527. |
| [8] | 李捷, 陈莹莹, 乔福云, 郅堤港, 郭正刚. 高原鼠兔干扰对高寒草甸β多样性的影响[J]. 植物生态学报, 2021, 45(5): 476-486. |
| [9] | 杨德春, 胡雷, 宋小艳, 王长庭. 降雨变化对高寒草甸不同植物功能群凋落物质量及其分解的影响[J]. 植物生态学报, 2021, 45(12): 1314-1328. |
| [10] | 王银柳, 耿倩倩, 黄建辉, 王常慧, 李磊, 哈斯木其尔, 牛国祥. 氮肥和种植密度对达乌里胡枝子的生长与生物固氮的影响[J]. 植物生态学报, 2021, 45(1): 13-22. |
| [11] | 陈锦, 宋明华, 李以康. 13C脉冲标记揭示放牧对高寒草甸同化碳分配的影响[J]. 植物生态学报, 2019, 43(7): 576-584. |
| [12] | 李全弟, 刘旻霞, 夏素娟, 南笑宁, 蒋晓轩. 甘南高寒草甸群落的物种-多度关系沿坡向的变化[J]. 植物生态学报, 2019, 43(5): 418-426. |
| [13] | 耿晓东, 旭日, 刘永稳. 青藏高原纳木错高寒草甸生态系统碳交换对多梯度增水的响应[J]. 植物生态学报, 2018, 42(3): 397-405. |
| [14] | 陈宁, 张扬建, 朱军涛, 李军祥, 刘瑶杰, 俎佳星, 丛楠, 黄珂, 王荔. 高寒草甸退化过程中群落生产力和物种多样性的非线性响应机制研究[J]. 植物生态学报, 2018, 42(1): 50-65. |
| [15] | 孙小妹, 陈菁菁, 李金霞, 李良, 韩国君, 陈年来. 高寒草甸植物化学计量比对磷添加的分层响应[J]. 植物生态学报, 2018, 42(1): 78-85. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19