
植物生态学报 ›› 2016, Vol. 40 ›› Issue (9): 861-870.DOI: 10.17521/cjpe.2015.0400
所属专题: 生物多样性
• 研究论文 • 下一篇
李艳朋1, 许涵1,*( ), 李意德1, 骆土寿1, 陈德祥1, 周璋1, 林明献2, 杨怀2
), 李意德1, 骆土寿1, 陈德祥1, 周璋1, 林明献2, 杨怀2
收稿日期:2015-11-10
接受日期:2016-07-23
出版日期:2016-09-10
发布日期:2016-09-29
通讯作者:
许涵
基金资助:
Yan-Peng LI1, Han XU1,*(), Yi-De LI1, Tu-Shou LUO1, De-Xiang CHEN1, Zhang ZHOU1, Ming-Xian LIN2, Huai YANG2
Received:2015-11-10
Accepted:2016-07-23
Online:2016-09-10
Published:2016-09-29
Contact:
Han XU
摘要:
物种多样性的空间分布格局和维持机制是群落生态学的基本问题。为了探讨海南尖峰岭地区物种多样性空间分布格局的尺度效应, 以海南尖峰岭热带山地雨林60 hm2样地为研究对象, 分析了物种丰富度、物种多度、Shannon-Wiener指数、Simpson指数以及Pielou均匀度指数随6个空间取样尺度(5 m × 5 m、10 m × 10 m、20 m × 20 m、40 m × 40 m、100 m ×100 m、200 m × 200 m)的变化。结果表明: 相比Simpson指数和Pielou均匀度指数, 物种丰富度、多度以及Shannon-Wiener指数具有更为明显的空间尺度效应; 物种丰富度的方差随取样尺度增加呈现单峰分布特征, 并且在20 m × 20 m尺度上达到最大值, 而物种多度的方差随取样尺度的增加而增大; 物种丰富度和多度的正相关性随着取样尺度的增加逐渐减小甚至消失, 这可能与随取样尺度增加生境异质性增加有关; 取样尺度对3个物种多样性指数空间分布的影响可能与研究区域内稀有种的组成有关。
李艳朋, 许涵, 李意德, 骆土寿, 陈德祥, 周璋, 林明献, 杨怀. 海南尖峰岭热带山地雨林物种多样性空间分布格局的尺度效应. 植物生态学报, 2016, 40(9): 861-870. DOI: 10.17521/cjpe.2015.0400
Yan-Peng LI, Han XU, Yi-De LI, Tu-Shou LUO, De-Xiang CHEN, Zhang ZHOU, Ming-Xian LIN, Huai YANG. Scale-dependent spatial patterns of species diversity in the tropical montane rain forest in Jianfengling, Hainan Island, China. Chinese Journal of Plant Ecology, 2016, 40(9): 861-870. DOI: 10.17521/cjpe.2015.0400
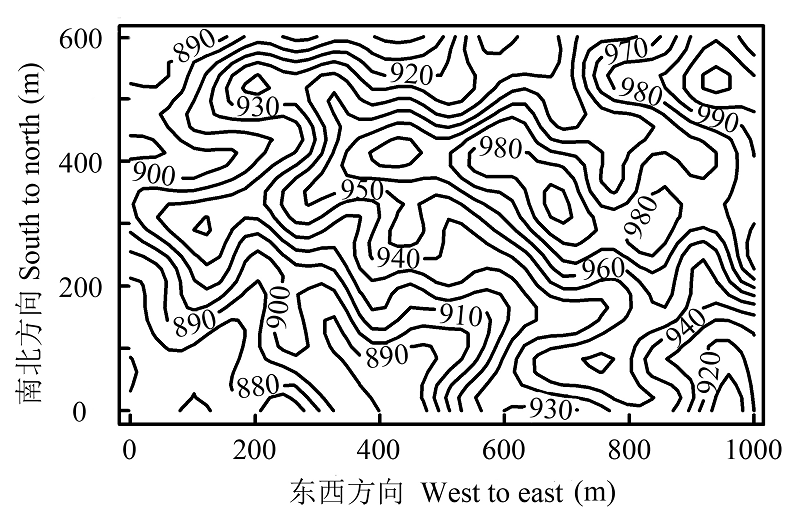
图1 海南尖峰岭60 hm2森林动态监测样地的地形图。
Fig. 1 Topography of the 60 hm2 Jianfengling Forest Dynamics Plot on Hainan Island.
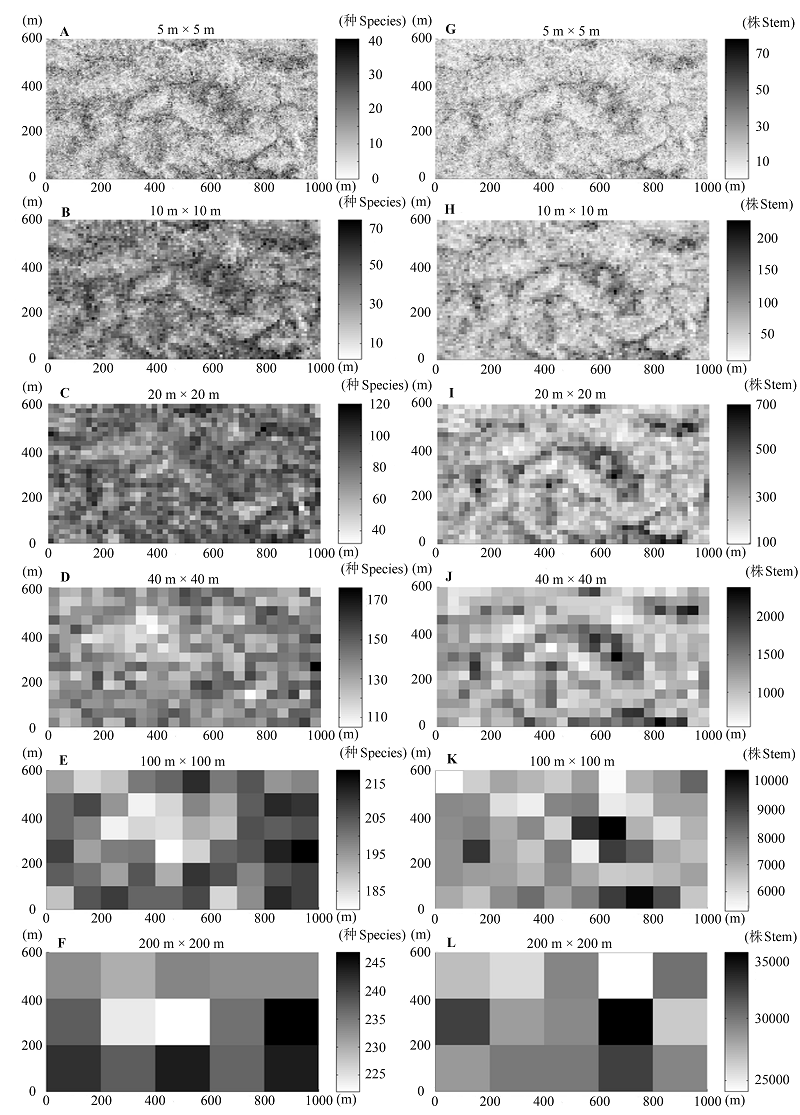
图2 物种丰富度(A-F)和多度(G-L)在不同尺度下的空间分布格局。标尺代表不同样方物种丰富度或多度的数值。
Fig. 2 Spatial distribution patterns of species richness (A-F) and abundance (G-L) at different scales. The sidebars represented the values of species richness or abundance at different plots, respectively.
| 取样尺度 Sampling scales | 5 m × 5 m | 10 m × 10 m | 20 m × 20 m | 40 m × 40 m | 100 m × 100 m | 200 m × 200 m |
|---|---|---|---|---|---|---|
| 相关系数 Correlation coefficients | 0.917 | 0.873 | 0.735 | 0.319 | -0.024 | 0.300 |
| 样方数 Number of quadrats | 24 000 | 6 000 | 1 500 | 375 | 60 | 15 |
| p | 0.000 | 0.000 | 0.000 | 0.000 | 0.856 | 0.277 |
表1 物种丰富度与物种多度的Spearman秩相关分析结果
Table 1 Spearman rank correlation coefficients between species richness and abundance at different spatial scales
| 取样尺度 Sampling scales | 5 m × 5 m | 10 m × 10 m | 20 m × 20 m | 40 m × 40 m | 100 m × 100 m | 200 m × 200 m |
|---|---|---|---|---|---|---|
| 相关系数 Correlation coefficients | 0.917 | 0.873 | 0.735 | 0.319 | -0.024 | 0.300 |
| 样方数 Number of quadrats | 24 000 | 6 000 | 1 500 | 375 | 60 | 15 |
| p | 0.000 | 0.000 | 0.000 | 0.000 | 0.856 | 0.277 |

图3 Shannon-Wiener指数(A-F)和Simpson指数(G-L)在不同尺度下的空间分布格局。标尺代表不同样方Shannon-Wiener指数或Simpson指数的数值。
Fig. 3 Spatial distribution patterns of Shannon-Wiener (A-F) and Simpson indices (G-L) at different scales. The sidebars represented the values of Shannon-Wiener or Simpson indices at different plots, respectively.
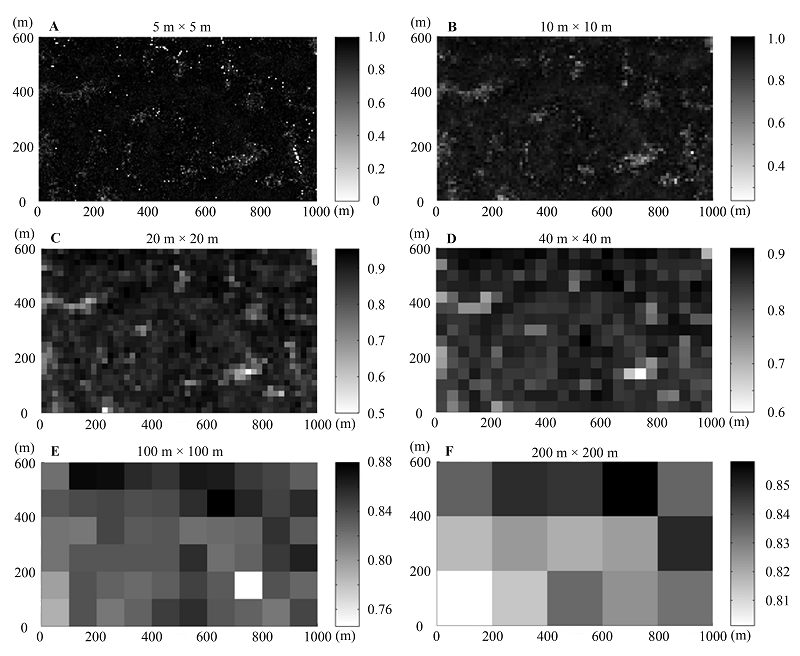
图4 Pielou均匀度指数在不同尺度下的空间分布格局。标尺代表不同样方Pielou均匀度指数的数值。
Fig. 4 Spatial distribution patterns of Pielou’s evenness index at different scales. The sidebars represented the values of Pielou’s evenness index at different plots.
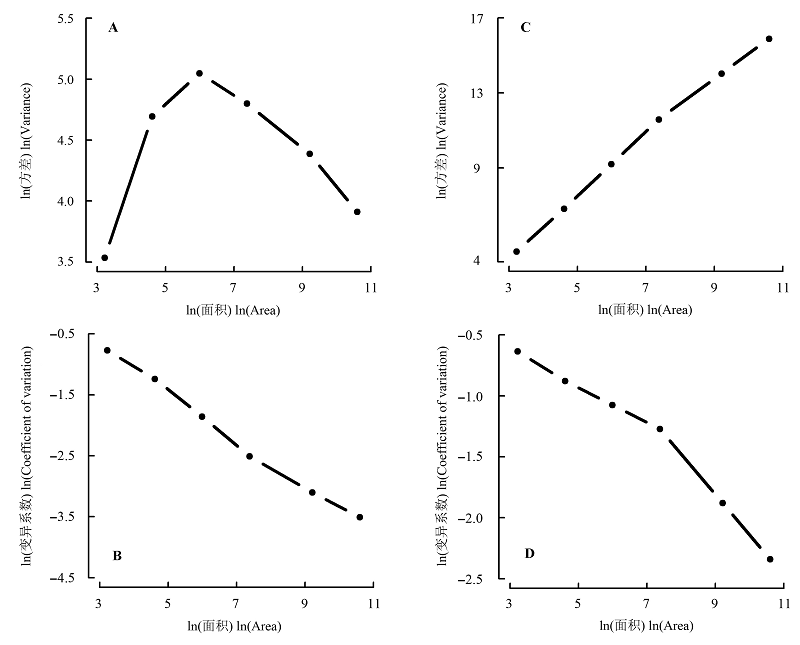
图5 物种丰富度(A, B)和多度(C, D)在不同尺度下的空间变异。
Fig. 5 Spatial variation of species richness (A, B) and abundance (C, D) at different scales.
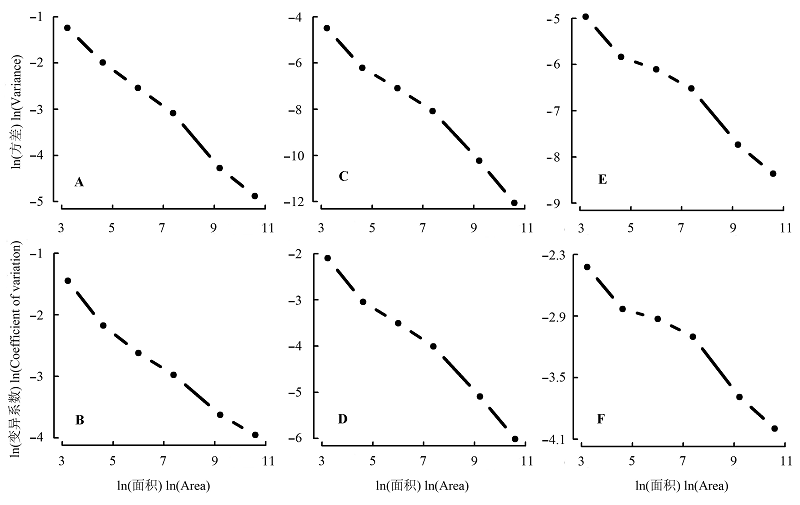
图6 Shannon-Wiener指数(A, B)、Simpson指数(C, D)以及Pielou均匀度指数(E, F)在不同尺度下的空间变异。
Fig. 6 Spatial variation of Shannon-Wiener (A, B), Simpson (C, D) and Pielou’s evenness indices (E, F) at different scales.
| 1 | Bartha S, Czárán T, Podani J (1998). Exploring plant community dynamics in abstract coenostate spaces.Abstracta Botanica, 22, 49-66. |
| 2 | Cheng JJ, Mi XC, Ma KP, Zhang JT (2011). Responses of species-abundance distribution to varying sampling scales in a subtropical broad-leaved forest.Biodiversity Science, 19, 168-177. (in Chinese with English abstract)[程佳佳, 米湘成, 马克平, 张金屯 (2011). 亚热带常绿阔叶林群落物种多度分布格局. 生物多样性, 19, 168-177.] |
| 3 | Colwell RK, Chao A, Gotelli NJ, Lin SY, Mao CX, Chazdon RL, Longino JT (2012). Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages.Journal of Plant Ecology, 5, 3-21. |
| 4 | Condit R (1998). Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. Springer, Berlin. |
| 5 | Crawley MJ, Harral JE (2001). Scale dependence in plant biodiversity.Science, 291, 864-868. |
| 6 | Fortin MJ, Payette S, Marineau K (1999). Spatial vegetation diversity index along a postfire successional gradient in the northern boreal forest.Ecoscience, 6, 204-213. |
| 7 | Giladi I, Ziv Y, May F, Jeltsch F (2011). Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem.Journal of Vegetation Science, 22, 983-996. |
| 8 | Harte J, Smith AB, Storch D (2009). Biodiversity scales from plots to biomes with a universal species-area curve.Ecology Letters, 12, 789-797. |
| 9 | He FL, LaFrankie JV, Song B (2002). Scale dependence of tree abundance and richness in a tropical rain forest, Malaysia.Landscape Ecology, 17, 559-568. |
| 10 | He FL, Legendre P (1996). On species-area relations.The American Naturalist, 148, 719-737. |
| 11 | He FL, Legendre P, Bellehumeur C, LaFrankie JV (1994). Diversity pattern and spatial scale: A study of a tropical rain forest of Malaysia.Environmental and Ecological Statistics, 1, 265-286. |
| 12 | Hill MO (1973). Diversity and evenness: A unifying notation and its consequences.Ecology, 54, 427-432. |
| 13 | Horne JK, Schneider DC (1995). Spatial variance in ecology.Oikos, 74, 18-26. |
| 14 | Hurlbert AH, Jetz W (2007). Species richness, hotspots, and the scale dependence of range maps in ecology and conserva- tion.Proceedings of the National Academy of Sciences of the United States of America, 14, 13384-13389. |
| 15 | Jiang YX, L JP (1991). Tropical Forest Ecosystem of Jian- fengling Mountain in Hainan Island, China. Science Press, Beijing. (in Chinese)[蒋有绪, 卢俊培 (1991). 中国海南岛尖峰岭热带林生态系统. 科学出版社, 北京.] |
| 16 | Juhász-Nagy P, Podani J (1983). Information theory methods for the study of spatial processes and succession.Plant Ecology, 51, 129-140. |
| 17 | Kallimanis AS, Halley JM, Vokou D, Sgardelis SP (2008). The scale of analysis determines the spatial pattern of woody species diversity in the Mediterranean environment.Plant Ecology, 196, 143-151. |
| 18 | Li YD, Chen BF, Zhou GY, Wu ZM, Zeng QB, Luo TS, Huang SN, Xie MD, Huang Q (2002). Research and Conservation of Tropical Forest and the Biodiversity: A Special Reference to Hainan Island, China. China Forestry Publishing House, Beijing. 47-62. (in Chinese)[李意德, 陈步峰, 周光益, 吴仲民, 曾庆波, 骆土寿, 黄世能, 谢明东, 黄全 (2002). 中国海南岛热带森林及其生物多样性保护研究. 中国林业出版社, 北京. 47-62.] |
| 19 | Li YD, Xu H, Luo TS, Chen DX, Lin MX (2012). Permanent Monitoring and Research Dataset of Chinese Ecosystem: Forest Ecosystem: Jianfengling Station (Bio-species Checklist). China Agriculture Press, Beijing. 10-35. (in Chinese)[李意德, 许涵, 骆土寿, 陈德祥, 林明献 (2012). 中国生态系统定位观测与研究数据集: 森林生态系统卷: 海南尖峰岭站(生物物种数据集). 中国农业出版社, 北京. 10-35.] |
| 20 | Ma KP, Huang JH, Yu SL, Chen LZ (1995). Plant community diversity in Dongling Mountain, Beijing, China: II, species richness, evenness and species diversities.Acta Ecologica Sinica, 15, 268-277. (in Chinese with English abstract)[马克平, 黄建辉, 于顺利, 陈灵芝 (1995). 北京东灵山地区植物群落多样性的研究: II丰富度、均匀度和物种多样性. 生态学报, 15, 268-277.] |
| 21 | Mouillot D, Leprêtre A (1999). A comparison of species diversity estimators.Researches on Population Ecology, 41, 203-215. |
| 22 | Peng Y, Qing FT, Mi K, Xue DY (2015). Study progress on spatial scale effects and coupling relationships of different levels in biodiversity.Acta Ecologica Sinica, 35, 577-583. (in Chinese with English abstract)[彭羽, 卿凤婷, 米凯, 薛达元 (2015). 生物多样性不同层次尺度效应及其耦合关系研究进展. 生态学报, 35, 577-583.] |
| 23 | Preston FW (1962). The canonical distribution of commonness and rarity: Part I.Ecology, 43, 185-215. |
| 24 | Rahbek C (2005). The role of spatial scale and the perception of large-scale species-richness patterns.Ecology Letters, 8, 224-239. |
| 25 | Ray C, Hastings A (1996). Density dependence: Are we searching at the wrong spatial scale?Journal of Animal Ecology, 65, 556-566. |
| 26 | Shi LL (2012). Study on the Spatial Heterogeneity of Soil Physical and Chemical Properties of Primary Tropical Montane Rainforest in Jianfengling, Hainan Island. Master degree dissertation, Chinese Academy of Forestry, Beijing. 26-41. (in Chinese with English abstract)[时雷雷 (2012). 海南尖峰岭热带山地雨林原始林土壤理化性质空间异质性研究. 硕士学位论文, 中国林业科学研究院, 北京. 26-41.] |
| 27 | Storch D, Marquet PA, Brown JH (2007). Scaling Biodiversity. Cambridge University Press, Cambridge, UK. 347-375. |
| 28 | Turner WR, Tjørve E (2005). Scale-dependence in species-area relationships.Ecography, 28, 721-730. |
| 29 | Wang XG, Hao ZQ, Ye J, Li BH, Yao XL (2008). Spatial variation of species diversity across scale in an old-growth temperate forest of China.Ecological Research, 23, 709-717. |
| 30 | Wang ZG, Ye WH, Cao HL, Lian JY (2008). Spatial distribu- tion of species diversity indices in a monsoon evergreen broadleaved forest at Dinghushan Mountain.Biodiversity Science, 16, 454-461. (in Chinese with English abstract)[王志高, 叶万辉, 曹洪麟, 练琚愉 (2008). 鼎湖山季风常绿阔叶林物种多样性指数空间分布特征. 生物多样性, 16, 454-461.] |
| 31 | Willis KJ, Whittaker RJ (2002). Species diversity-scale matters.Science, 295, 1245-1248. |
| 32 | Wilson JB, Steel JB, King WM, Gitay H (1999). The effect of spatial scale on evenness.Journal of Vegetation Science, 10, 463-468. |
| 33 | Xu H, Li YD, Luo TS, Chen DX, Lin MX, Wu JH, Li YP, Yang H, Zhou Z (2015). Jianfengling Tropical Mountain Rain Forest Dynamic Plot: Community Characteristic, Tree Species and Their Distribution Patterns. China Forestry Publishing House, Beijing. 34-48. (in Chinese)[许涵, 李意德, 骆土寿, 陈德祥, 林明献, 吴建辉, 李艳朋, 杨怀, 周璋 (2015). 海南尖峰岭热带山地雨林——群落特征、树种及其分布格局. 中国林业出版社, 北京. 34-48.] |
| 34 | Xu H, Liu SR, Li YD, Zang RG, He FL (2012). Assessing non-parametric and area-based methods for estimating regional species richness.Journal of Vegetation Science, 23, 1006-1012. |
| 35 | Xu Q, Zhang F, Xu ZQ, Jia YL, You JM (2011). Some characteristics of Simpson index and the Shannon-Wiener index and their dilution effect.Pratacultural Science, 4, 527-531. (in Chinese with English abstract)[许晴, 张放, 许中旗, 贾彦龙, 尤建民 (2011). Simpson指数和Shannon-Wiener指数若干特征的分析及“稀释效应”. 草业科学, 4, 527-531.] |
| 36 | Ye WH, Cao HL, Huang ZL, Lian JY, Wang ZG, Li L, Wei SG, Wang ZM (2008). Community structure of a 20 hm2 lower subtropical evergreen broadleaved forest plot in Dinghushan, China. Journal of Plant Ecology (Chinese Version), 32, 274-286. (in Chinese with English abstract)[叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明 (2008). 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 32, 274-286.] |
| 37 | Zhang ZH, Hu G, Zhu JD, Ni J (2012). Scale-dependent spatial variation of species abundance and richness in two mixed evergreen-deciduous broad-leaved karst forests, Southwest China.Acta Ecologica Sinica, 32, 5663-5672. (in Chinese with English abstract)[张忠华, 胡刚, 祝介东, 倪健 (2012). 喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应. 生态学报, 32, 5663-5672.] |
| [1] | 牛一迪, 蔡体久. 大兴安岭北部次生林演替过程中物种多样性的变化及其影响因子[J]. 植物生态学报, 2024, 48(3): 349-363. |
| [2] | 李娜, 唐士明, 郭建英, 田茹, 王姗, 胡冰, 罗永红, 徐柱文. 放牧对内蒙古草地植物群落特征影响的meta分析[J]. 植物生态学报, 2023, 47(9): 1256-1269. |
| [3] | 杨鑫, 任明迅. 环南海区域红树物种多样性分布格局及其形成机制[J]. 植物生态学报, 2023, 47(8): 1105-1115. |
| [4] | 于笑, 纪若璇, 任天梦, 夏新莉, 尹伟伦, 刘超. 中国北方蒙古莸群落的分布、特征和分类[J]. 植物生态学报, 2023, 47(8): 1182-1192. |
| [5] | 钟姣, 姜超, 刘世荣, 龙文兴, 孙建新. 海南长臂猿食源植物的潜在物种丰富度分布格局[J]. 植物生态学报, 2023, 47(4): 491-505. |
| [6] | 李晓田, 王铁娟, 韩文娟, 张丽, 张慧, 刘晓婷, 刘雅洁. 东阿拉善珍稀濒危植物绵刺种群结构与点格局分析[J]. 植物生态学报, 2023, 47(4): 506-514. |
| [7] | 朱华, 谭运洪. 中国热带雨林的群落特征、研究现状及问题[J]. 植物生态学报, 2023, 47(4): 447-468. |
| [8] | 杨元合, 张典业, 魏斌, 刘洋, 冯雪徽, 毛超, 徐玮婕, 贺美, 王璐, 郑志虎, 王媛媛, 陈蕾伊, 彭云峰. 草地群落多样性和生态系统碳氮循环对氮输入的非线性响应及其机制[J]. 植物生态学报, 2023, 47(1): 1-24. |
| [9] | 董六文, 任正炜, 张蕊, 谢晨笛, 周小龙. 功能多样性比物种多样性更好解释氮添加对高寒草地生物量的影响[J]. 植物生态学报, 2022, 46(8): 871-881. |
| [10] | 曾凯娜, 孙浩然, 申益春, 任明迅. 海南羊山湿地的传粉网络及其季节动态[J]. 植物生态学报, 2022, 46(7): 775-784. |
| [11] | 彭鑫, 金光泽. 植物特性和环境因子对阔叶红松林暗多样性的影响[J]. 植物生态学报, 2022, 46(6): 656-666. |
| [12] | 陈丽, 田新民, 任正炜, 董六文, 谢晨笛, 周小龙. 养分添加对天山高寒草地植物多样性和地上生物量的影响[J]. 植物生态学报, 2022, 46(3): 280-289. |
| [13] | 郝建锋, 周润惠, 姚小兰, 喻静, 陈聪琳, 向琳, 王姚瑶, 苏天成, 齐锦秋. 二代野猪放牧对夹金山针阔混交林物种多样性与土壤理化性质的影响[J]. 植物生态学报, 2022, 46(2): 197-207. |
| [14] | 张义, 程杰, 苏纪帅, 程积民. 长期封育演替下典型草原植物群落生产力与多样性关系[J]. 植物生态学报, 2022, 46(2): 176-187. |
| [15] | 宋语涵, 张鹏, 金光泽. 阔叶红松林不同演替阶段灌木叶片碳氮磷化学计量特征及其影响因素[J]. 植物生态学报, 2021, 45(9): 952-960. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19