
植物生态学报 ›› 2018, Vol. 42 ›› Issue (2): 209-219.DOI: 10.17521/cjpe.2017.0132
彭曦,闫文德,王凤琪,王光军,玉昉永,赵梅芳( )
)
出版日期:2018-02-20
发布日期:2018-04-16
通讯作者:
赵梅芳
基金资助:
PENG Xi,YAN Wen-De,WANG Feng-Qi,WANG Guang-Jun,YU Fang-Yong,ZHAO Mei-Fang()
Online:2018-02-20
Published:2018-04-16
Contact:
Mei-Fang ZHAO
Supported by:摘要:
随着叶片功能性状研究的不断深入, 通过简单易测量的叶片指标, 同时探究植物生活史权衡对策和估算林分生产力的研究需求日益增长, 例如叶干质量比(LDMC)和比叶面积(SLA)的相互转换。杉木(Cunninghamia lanceolata)是亚热带重要的常绿针叶树种, 基于LDMC对杉木SLA进行估算, 能够为核算SLA提供途径, 为机理解释和生产估算构建连接途径, 为小区域到大尺度、精算到估算搭建桥梁。该研究在湖南会同和河南信阳两个杉木生长区, 对处于不同小生境(坡向、坡位和冠层深度)以及不同生活史(林龄和叶龄)的叶片进行抽样和采集, 通过测得不同叶龄的单叶LDMC和SLA, 初步探究在不同因子下两个性状值的分布差异, 进一步基于LDMC构建SLA估算模型并讨论以叶龄为差分因子对模型的影响。结果表明: 1)杉木SLA平均值为(103.15 ± 69.54) cm 2·g -1, LDMC为0.39 ± 0.11; 2)杉木LDMC和SLA可用非线性模型进行估算, 模型符合估算要求; 3)其中一年生叶的拟合效果最好, 老叶(大于二年生叶)的拟合优度较低, 老叶较低的SLA (52.28-75.74 cm 2·g -1)可能暗示LDMC的变化保持相对独立性。该研究基于杉木LDMC的SLA估算模型可信且有效, 且不同叶龄对LDMC和SLA的影响可能预示着杉木叶片的响应敏感性和生活史权衡策略。
彭曦, 闫文德, 王凤琪, 王光军, 玉昉永, 赵梅芳. 基于叶干质量比的杉木比叶面积估算模型的构建. 植物生态学报, 2018, 42(2): 209-219. DOI: 10.17521/cjpe.2017.0132
PENG Xi, YAN Wen-De, WANG Feng-Qi, WANG Guang-Jun, YU Fang-Yong, ZHAO Mei-Fang. Specific leaf area estimation model building based on leaf dry matter content of Cunninghamia lanceolata. Chinese Journal of Plant Ecology, 2018, 42(2): 209-219. DOI: 10.17521/cjpe.2017.0132
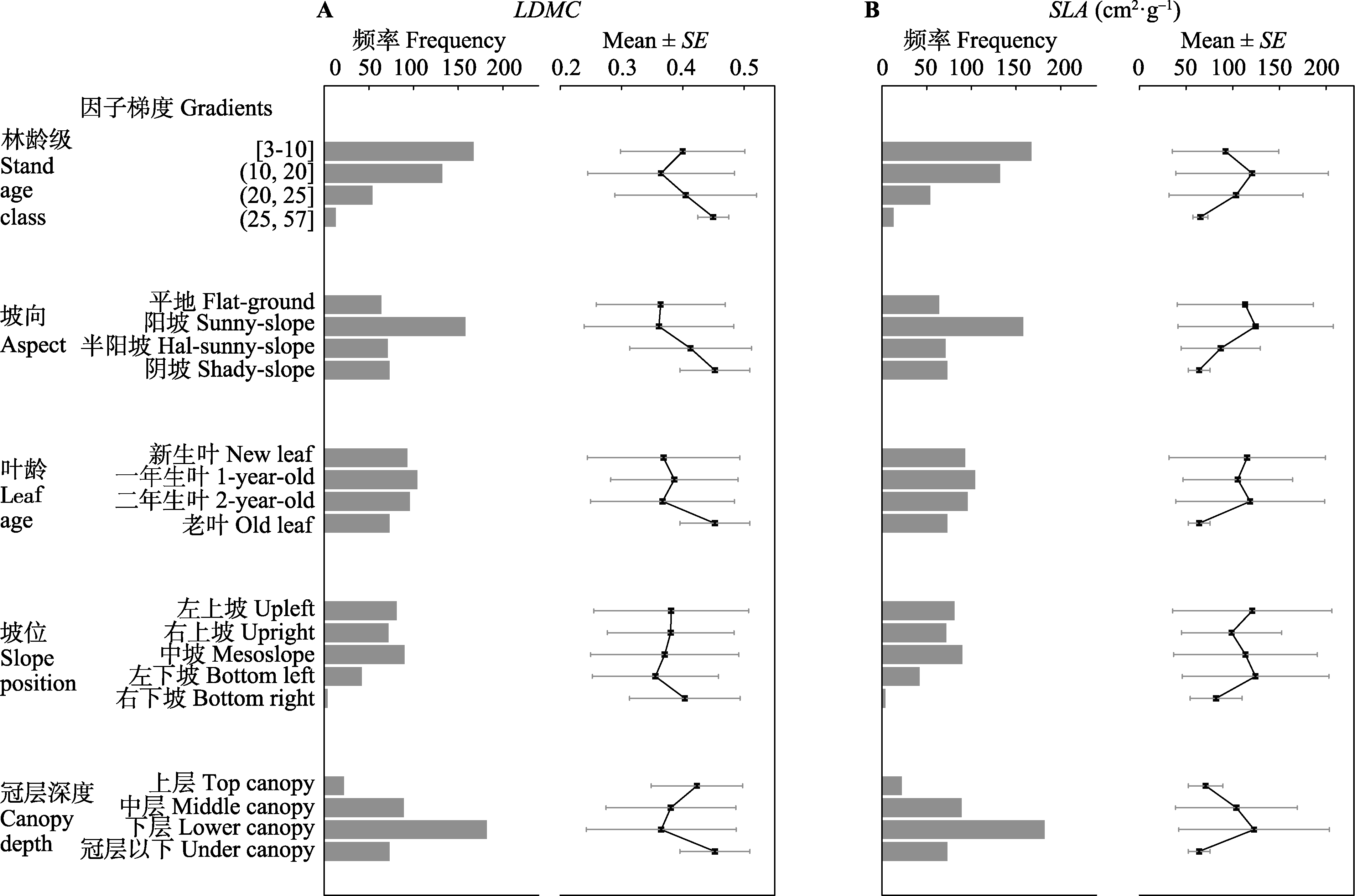
图1 杉木叶干质量比(LDMC) (A)、比叶面积(SLA) (B)在不同因子变异梯度下的总体抽样特征及均值离散化程度(n = 374)。
Fig. 1 Summary characteristics and degree of discretization of leaf dry matter content (LDMC) (A) and specific leaf area (SLA) (B) of Cunninghamia lanceolata along different factors gradients (n = 374).
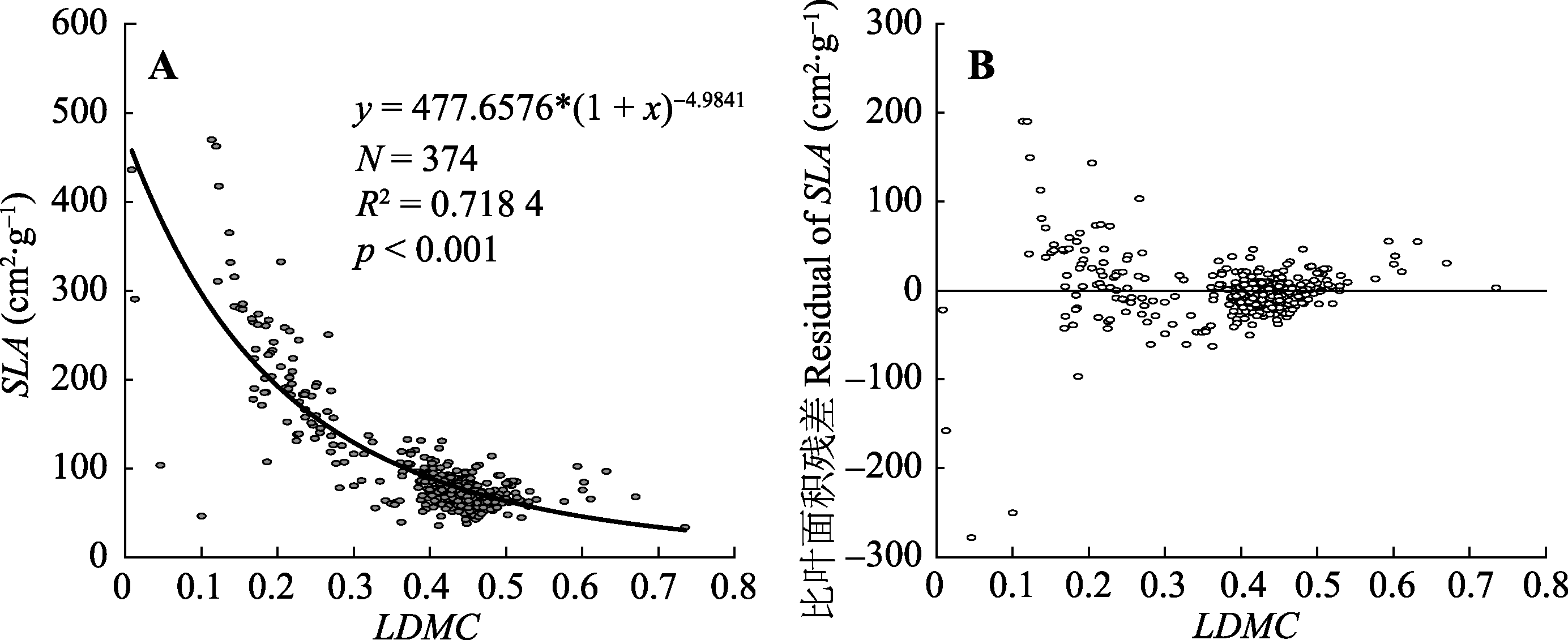
图2 杉木基于叶干质量比(LDMC)的比叶面积(SLA)非线性拟合模型(A)和SLA对LDMC的回归残差散点图(B)。模型选择y = a(1 + x)b; N, 样本量; R2, 修正的决定系数; p, 差异性显著度。
Fig. 2 Nonlinear fitting model of specific leaf area (SLA) based on leaf dry matter content (LDMC) (A) and SLA scatter plot the regression residuals of LDMC (B) of Cunninghamia lanceolata. Model selection y = a(1 + x)b; N, sampling size; R2, adjusted R-squared; p, significant difference degree.
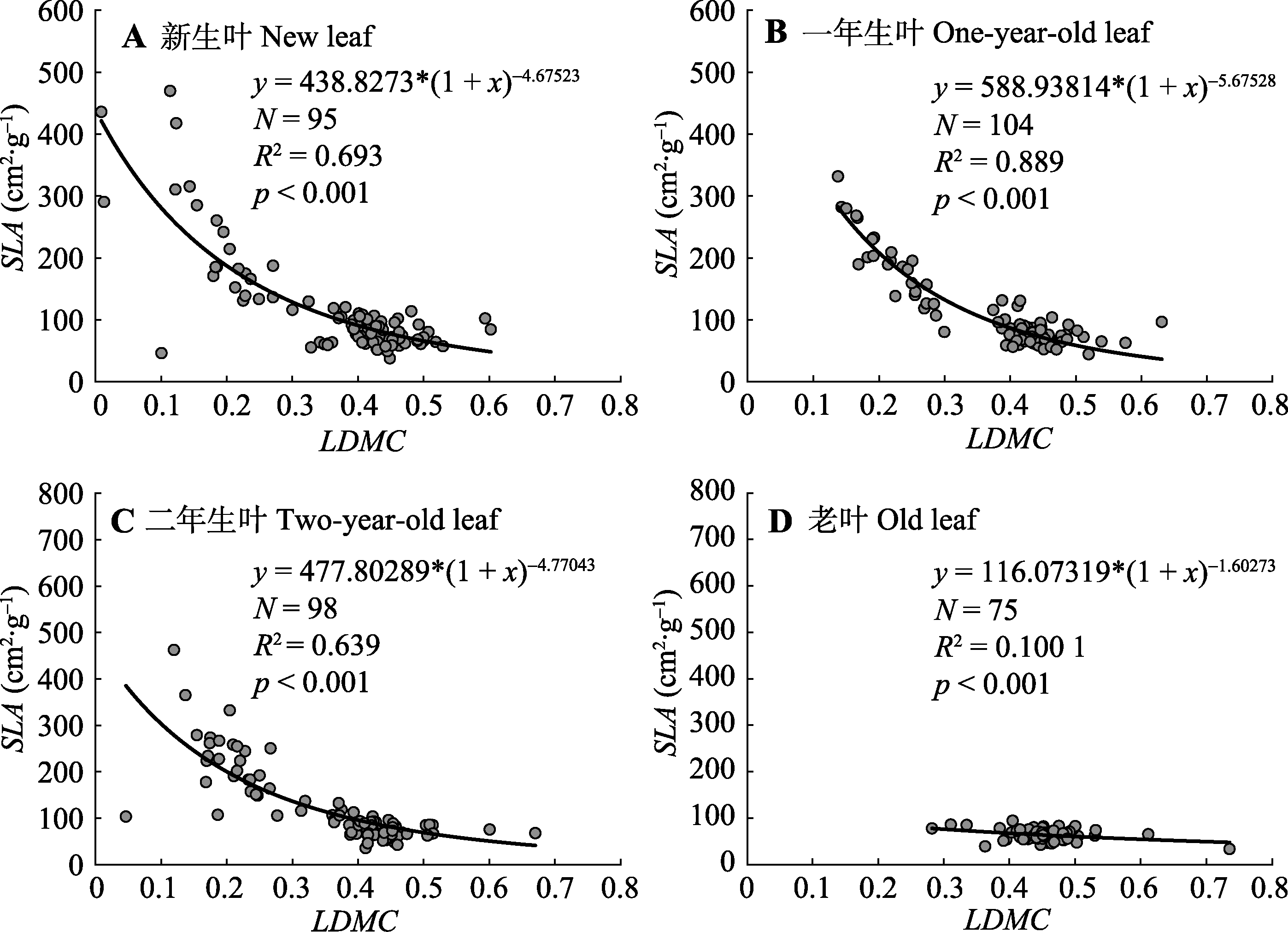
图3 不同叶龄梯度下杉木基于叶干质量比(LDMC)的比叶面积(SLA)非线性拟合模型。模型选择y = a(1 + x)b; N, 样本量; R2, 修正的决定系数; p, 差异性显著度。
Fig. 3 Nonlinear fitting model of specific leaf area (SLA) based on leaf dry matter content (LDMC) of Cunninghamia lanceolata along different leaf age gradients. Model selection y = a(1 + x)b; N, sampling size; R2, adjusted R-squared; p, significant difference degree.
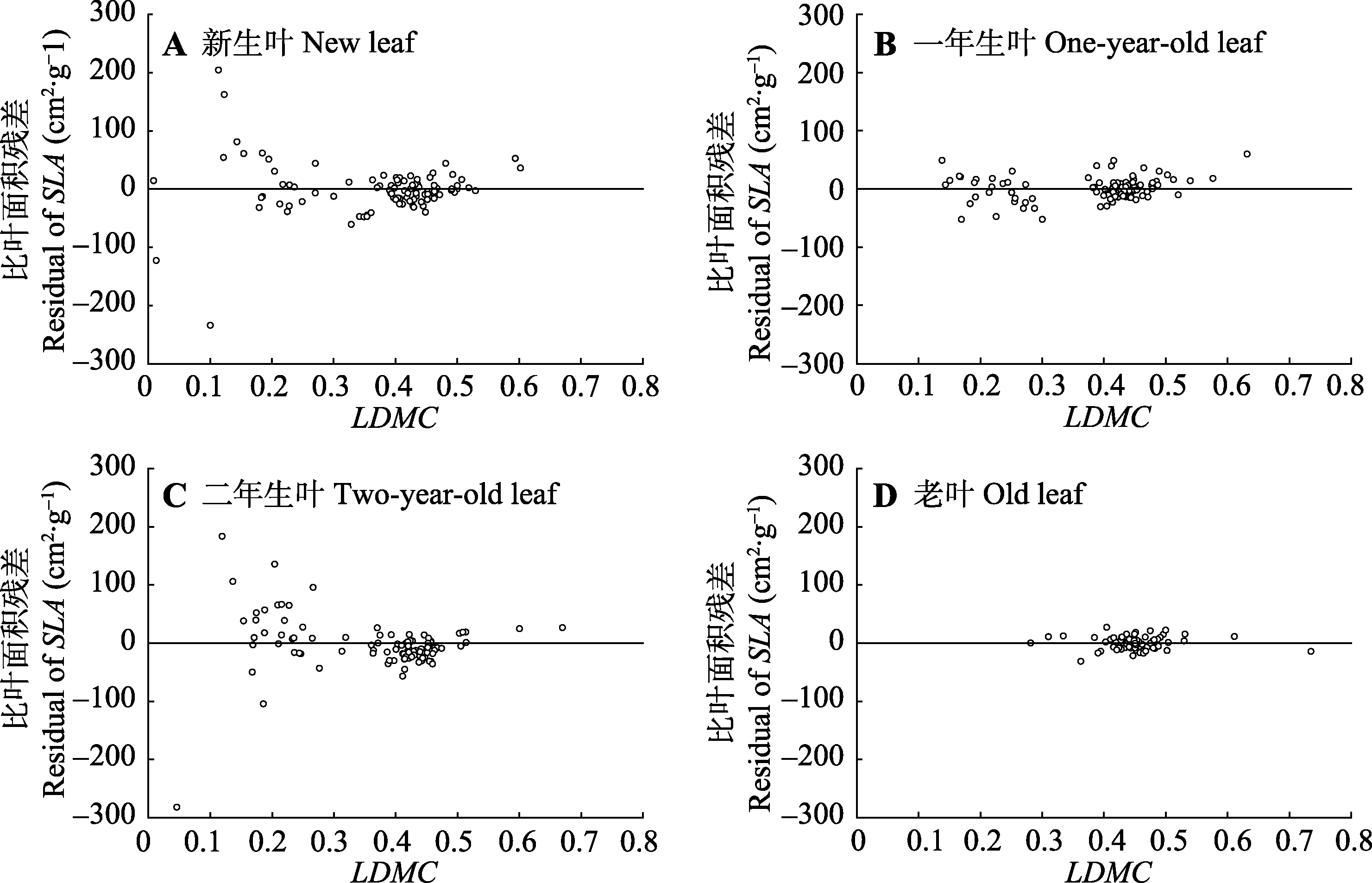
图4 不同叶龄梯度下杉木比叶面积(SLA)对叶干质量比(LDMC)的回归残差散点图。
Fig. 4 Specific leaf area (SLA) scatter plot the regression residuals of leaf dry matter content (LDMC) of Cunninghamia lanceolata along different leaf age gradients.
| [1] |
Athokpam FD, Garkoti SC, Borah N ( 2013). Periodicity of leaf growth and leaf dry mass changes in the evergreen and deciduous species of southern Assam, India. Ecological Research, 29, 153-165.
DOI URL |
| [2] |
Bertin N ( 1998). Short and long term fluctuations of the leaf mass per area of tomato plants—Implications for growth models. Annals of Botany, 82, 71-81.
DOI URL |
| [3] |
Cantón Y, Del Barrio G, Solé-Benet A, Lázaro R ( 2004). Topographic controls on the spatial distribution of ground cover in the tabernas badlands of SE Spain. Catena, 55, 341-365.
DOI URL |
| [4] |
Cornelissen JHC, Lavorel S, Garnier E ( 2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 51, 335-380.
DOI URL |
| [5] | Dang JJ, Zhao CZ, Li Y, Hou ZJ, Dong XG ( 2015). Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China. Chinese Journal of Plant Ecology, 39, 23-31. |
| [ 党晶晶, 赵成章, 李钰, 侯兆疆, 董小刚 ( 2015). 祁连山高寒草地甘肃臭草叶性状与坡向间的关系. 植物生态学报, 39, 23-31.] | |
| [6] |
Díaz S, Cabido M ( 2009). Plant functional types and ecosystem function in relation to global change. Journal of Vegetation Science, 8, 463-474.
DOI URL |
| [7] |
Díaz S, Cabido M, Fernando C ( 1998). Plant functional traits and environmental filters at a regional scale. Journal of Vegetation Science, 9, 113-122.
DOI URL |
| [8] | Duan YY, Song LJ, Niu SQ, Huang T, Yang GH, Hao WF ( 2017). Variation of leaf functional traits of different aged Robinia pseudoacacia communities and relationships with soil nutrients. Chinese Journal of Applied Ecology, 28, 28-36. |
| [ 段媛媛, 宋丽娟, 牛素旗, 黄婷, 杨改河, 郝文芳 ( 2017). 不同林龄刺槐叶功能性状差异及其与土壤养分的关系. 应用生态学报, 28, 28-36.] | |
| [9] |
Ellsworth DS, Reich PB ( 1993). Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia, 96, 169-178.
DOI URL PMID |
| [10] |
Field C, Mooney HAJ ( 1983). Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a california shrub. Oecologia, 56, 348-355.
DOI URL |
| [11] |
Frak E, Roux XL, Millard P, Dreyed E, Jaouen G, Saint-Joanis B, Wendler R ( 2001). Changes in total leaf nitrogen and partitioning of leaf nitrogen drive photosynthetic acclimation to light in fully developed walnut leaves. Plant, Cell & Environment, 24, 1279-1288.
DOI URL |
| [12] |
Gao JL, Luo FM, Zhao YM, Zhang JB, Yuan WJ, Sun F, Hao YG ( 2016). Specific leaf area and leaf dry matter content of three kinds of poplar in Ulan Buh desert oasis. Journal of Northwest Forestry University, 31(1), 15-20.
DOI URL |
|
[ 高君亮, 罗凤敏, 赵英铭, 张景波, 原伟杰, 孙非, 郝玉光 ( 2016). 乌兰布和沙漠绿洲3种杨树比叶面积和叶干物质含量研究. 西北林学院学报, 31(1), 15-20.]
DOI URL |
|
| [13] |
Garnier E, Cordonnier P, Guillerm JL ( 1997). Specific leaf area and leaf nitrogen concentration in annual and perennial grass species growing in mediterranean old-fields. Oecologia, 111, 490-498.
DOI URL PMID |
| [14] |
Garnier E, Laurent G, Bellmann A, Debain S, Berthelier P, Ducout B, Roumet C, Navas ML ( 2001 a). Consistency of species ranking based on functional leaf traits. New Phytologist, 152, 69-83.
DOI URL |
| [15] |
Garnier E, Shipley B, Roumet C, Laurent G ( 2001 b). A standardized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology, 15, 688-695.
DOI URL |
| [16] |
Gratani L, Bombelli A ( 2000). Correlation between leaf age and other leaf traits in three mediterranean maquis shrub species:Quercus ilex, Phillyrea latifolia and Cistus incanus. Environmental and Experimental Botany, 43, 141-153.
DOI URL |
| [17] |
Greenwood MS, Ward MH, Day ME, Adams SL, Bond BJ ( 2008). Age-related trends in red spruce foliar plasticity in relation to declining productivity. Tree Physiology, 28, 225-232.
DOI URL |
| [18] |
Grime JP, Thompson K, Hunt R, Hodgson JG, Cornelissen JHC, Rorison IH, Hendry GAF, Ashenden TW, Askew AP, Band SR, Booth RE, Bossard CC, Campbell BD, Cooper JEL, Davison AW, Gupta PL, Hall W, Hand DW, Hannah MA, Hillier SH, Hodkinson DJ, Jalili A, Liu Z, Mackey JML, Matthews N, Mowforth MA, Neal AM, Reader RJ, Reiling K, Ross-Fraser W, Spencer RE, Sutton F, Tasker DE, Thorpe PC, Whitehouse J ( 1997). Integrated screening validates primary axes of specialisation in plants. Oikos, 79, 259-281.
DOI URL |
| [19] |
Hodgson JG, Montserrat-Martí G, Charles M, Jones G, Wilson P, Shipley B, Sharafi M, Cerabolini BEL, Cornelissen JHC, Band SR, Bogard A, Castro-Díez P, Guerrero- Campo J, Palmer C, Pérez-Rontomé MC, Carter G, Hynd A, Romo-Díez A, de Torres Espuny L, Royo Pla F ( 2011). Is leaf dry matter content a better predictor of soil fertility than specific leaf area? Annals of Botany, 108, 1337-1345.
DOI URL PMID |
| [20] |
Hou ZJ, Zhao CZ, Li Y, Zhang Q, Ma XL ( 2014). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland. Chinese Journal of Plant Ecology, 38, 281-288.
DOI URL |
|
[ 侯兆疆, 赵成章, 李钰, 张茜, 马小丽 ( 2014). 不同坡向高寒退化草地狼毒株高和枝条数的权衡关系. 植物生态学报, 38, 281-288.]
DOI URL |
|
| [21] |
Hu YS, Yao XY, Liu YH ( 2015). Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains. Acta Ecologica Sinica, 35, 1480-1487.
DOI URL |
|
[ 胡耀升, 么旭阳, 刘艳红 ( 2015). 长白山森林不同演替阶段比叶面积及其影响因子. 生态学报, 35, 1480-1487.]
DOI URL |
|
| [22] |
Huang HX, Yang XD, Sun BW, Zhang ZH, Yan ER ( 2014). Variability and association of leaf traits between current-?year and former-year leaves in evergreen trees in Tiantong, Zhejiang, China. Chinese Journal of Plant Ecology, 37, 912-921.
DOI URL |
|
[ 黄海侠, 杨晓东, 孙宝伟, 张志浩, 阎恩荣 ( 2014). 浙江天童常绿植物当年生与往年生叶片性状的变异与关联. 植物生态学报, 37, 912-921.]
DOI URL |
|
| [23] | Huo H, Wang CK ( 2007). Effects of canopy position and leaf age on photosynthesis and transpiration of Pinus koraiensis. Chinese Journal of Applied Ecology, 18, 1181-1186. |
| [ 霍宏, 王传宽 ( 2007). 冠层部位和叶龄对红松光合蒸腾特性的影响. 应用生态学报, 18, 1181-1186.] | |
| [24] | James SA, Bell DT ( 2000). Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. Globulus provenances. Tree Physiology, 20, 1007-1018. |
| [25] |
Jiang AP, Jiang JM, Liu J ( 2016). Response of leaf traits of Sassafras tsumu (Hemsl.) Hemsl.along an altitudinal gradient. . Chinese Journal of Ecology, 25, 1467-1474.
DOI URL |
|
[ 蒋艾平, 姜景民, 刘军 ( 2016). 檫木叶片性状沿海拔梯度的响应特征. 生态学杂志, 25, 1467-1474.]
DOI URL |
|
| [26] |
Jullien A, Allirand J-M, Mathieu A, Andrieu B, Ney B ( 2009). Variations in leaf mass per area according to N nutrition, plant age, and leaf position reflect ontogenetic plasticity in winter oilseed rape (Brassica napus l.). Field Crops Research, 114, 188-197.
DOI URL |
| [27] |
Karavin N ( 2013). Effects of leaf and plant age on specific leaf area in deciduous tree species Quercus cerris L. var. cerris. Bangladesh Journal of Botany, 42, 301-306.
DOI URL |
| [28] |
Kattge J, Díaz S, Lavorel S, Prentice IC, Leadley P, B?nisch G, Garnier E, Westoby M, Reich PB, Wright IJ, Cornelissen JHC, Violle C, Harrison SP, Van Bodegom PM, Reichstein M, Enquist BJ, Soudzilovskaia NA, Ackerly DD, Anand M, Atkin O, Bahn M, Baker TR, Baldocchi D, Bekker R, Blanco CC, Blonder B, Bond WJ, Bradstock R, Bunker DE, Casanoves F, Cavender-Bares J, Chambers JQ, Chapin III FS, Chave J, Coomes D, Cornwell WK, Craine JM, Dobrin BH, Duarte L, Durka W, Elser J, Esser G, Estiarte M, Fagan WF, Fang J, Fernández-Méndez F, Fidelis A, Finegan B, Flores O, Ford H, Frank D, Freschet GT, Fyllas NM, Gallagher RV, Green WA, Gutierrez AG, Hickler T, Higgins SI, Hodgson JG, Jalili A, Jansen S, Joly CA, Kerkhoff AJ, Kirkup D, Kitajima K, Kleyer M, Klotz S, Knops JMH, Kramer K, Kühn I, Kurokawa H, Laughlin D, Lee TD, Leishman M, Lens F, Lenz T, Lewis SL, Lloyd J, Llusià J, Louault F, Ma S, Mahecha MD, Manning P, Massad T, Medlyn BE, Messier J, Moles AT, Müller SC, Nadrowski K, Naeem S, Niinemets ü, N?llert S, Nüske A, Ogaya R, Oleksyn J, Onipchenko VG, Onoda Y, Ordo?Ez J, Overbeck G, Ozinga WA, Pati?o S, Paula S, Pausas JG, Pe?uelas J, Phillips OL, Pillar V, Poorter H, Poorter L, Poschlod P, Prinzing A, Proulx R, Rammig A, Reinsch S, Reu B, Sack L, Salgado-Negret B, Sardans J, Shiodera S, Shipley B, Siefert A, Sosinski E, Soussana JF, Swaine E, Swenson N, Thompson K, Thornton P, Waldram M, Weiher E, White M, White S, Wright SJ, Yguel B, Zaehle S, Zanne AE, Wirth C ( 2011). Try—A global database of plant traits. Global Change Biology, 17, 2905-2935.
DOI URL PMID |
| [29] |
Kattge J, Knorr W, Raddatz T, Wirth C ( 2009). Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Global Change Biology, 15, 976-991.
DOI URL |
| [30] | Kayama M, Sasa K, Koike T ( 2002). Needle life span, photosynthetic rate and nutrient concentration of Picea glehnii, P. jezoensis and P. abies planted on serpentine soil in northern Japan. Tree Physiology, 22, 707-716. |
| [31] |
Landsberg JJ, Gower ST ( 1997). Applications of physiological ecology to forest management. Tree Physiology, 17, 601-602.
DOI URL |
| [32] |
Lavorel S, Garnier E ( 2002). Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the holy grail. Functional Ecology, 16, 545-556.
DOI URL |
| [33] | Li K, Xiang WH ( 2011). Comparison of specific leaf area, SPAD value and seed mass among subtropical tree species in hilly area of central Hunan, China. Journal of Central South University of Forestry & Technology, 31, 213-218. |
| [ 李凯, 项文化 ( 2011). 湘中丘陵区12个树种比叶面积、SPAD值和种子干质量的比较. 中南林业科技大学学报, 31, 213-218.] | |
| [34] |
Li XR, Liu QJ, Cai Z, Ma ZQ ( 2007). Specific leaf area and leaf area index of conifer plantations in Qianyanzhou station of subtropical China. Journal of Plant Ecology (Chinese Version), 31, 93-101.
DOI URL |
|
[ 李轩然, 刘琪璟, 蔡哲, 马泽清 ( 2007). 千烟洲针叶林的比叶面积及叶面积指数. 植物生态学报, 31, 93-101.]
DOI URL |
|
| [35] |
Li YL, Cui JY, Su YZ ( 2005). Specific leaf area and leaf dry matter content of some plants in different dune habitats. Acta Ecologica Sinica, 25, 304-311.
DOI URL |
|
[ 李玉霖, 崔建垣, 苏永中 ( 2005). 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 25, 304-311.]
DOI URL |
|
| [36] | Liu ZM ( 2014). Research on the Nutrient Dynamics and Internal Absorption of Cunninghamia lanceolata Foliar. Master degree dissertation, Fujian Agriculture and Forestry University, Fuzhou. |
| [ 刘桌明 ( 2014). 杉木针叶养分含量动态及其内吸收率研究. 硕士学位论文, 福建农林大学, 福州.] | |
| [37] | Matsuzaki J, Norisada M, Kodaira J, Suzuki M, Tange T (2004). Shoots grafted into the upper crowns of tall Japanese cedar ( Cryptomeria japonica D. Don) show foliar gas exchange characteristics similar to those of intact shoots. Trees, 19, 198-203. |
| [38] |
Mencuccini M, Martínez-Vilalta J, Vanderklein D, Hamid HA, Korakaki E, Lee S, Michiels B ( 2005). Size-mediated ageing reduces vigour in trees. Ecology Letter, 8, 1183-1190.
DOI URL PMID |
| [39] | Miao YM, Lü JZ, Bi RC ( 2012). Distribution pattern of leaf traits in canopy of typical broad-leaved trees in Taiyue Mountain. Guihaia, 32, 483-486. |
| [ 苗艳明, 吕金枝, 毕润成 ( 2012). 太岳山典型阔叶乔木冠层叶片性状的分布格局. 广西植物, 32, 483-486.] | |
| [40] |
Nicotra AB, Leigh A, Boyce CK, Jones CS, Niklas KJ, Royer DL, Tsukaya H ( 2011). The evolution and functional significance of leaf shape in the angiosperms. Functional Plant Biology, 38, 535-552.
DOI URL |
| [41] |
Ono K, Nishi Y, Watanabe A, Terashima I ( 2001). Possible mechanisms of adaptive leaf senescence. Plant Biology, 3, 234-243.
DOI URL |
| [42] |
Osnas JL, Lichstein JW, Reich PB, Pacala SW ( 2013). Global leaf trait relationships: Mass, area, and the leaf economics spectrum. Science, 340, 741-744.
DOI URL |
| [43] | Ou XL, Liu YH ( 2017). Age, slope aspectsand diameter classes affect the leaf functional traits of Pinus tabulaeformis in Songshan, Beijing. Journal of Nanjing Forestry University (Natural Sciences Edition), 41(4), 80-88. |
| [ 欧晓岚, 刘艳红 ( 2017). 北京松山不同坡向和径级油松异龄叶功能性状特征. 南京林业大学学报(自然科学版), 41(4), 80-88.] | |
| [44] | Qin XH ( 2015). Spatial heterogeneity of specific leaf area and leaf construction cost of Quercus liaotungensis canopy. Forest Resources Management, ( 4), 145-150. |
| [ 覃鑫浩 ( 2015). 辽东栎冠层叶建成消耗与比叶面积的空间异质性. 林业资源管理, ( 4), 145-150.] | |
| [45] |
Reich PB, Uhl C, Walters MB, Ellsworth DS (1991a). Leaf lifespan as a determinant of leaf structure and function among 23 amazonian tree species. Oecologia, 86, 16-24.
DOI URL PMID |
| [46] | Reich PB, Walters MB, Ellsworth DS (1991 b). Leaf age and season influence the relationships between leaf nitrogen, leaf mass per area and photosynthesis in maple and oak trees. Plant, Cell & Environment, 14, 251-259. |
| [47] |
Reich PB, Walters MB, Ellsworth DS ( 1992). Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecological Monographs, 62, 365-392.
DOI URL |
| [48] |
Reich PB, Walters MB, Ellsworth DS, Vose JM, Volin JC, Gresham C, Bowman WD ( 1998). Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: A test across biomes and functional groups. Oecologia, 114, 471-482.
DOI URL PMID |
| [49] | Ren Y, Lu Q, Wu B, Li MH ( 2015). Specific leaf area and leaf dry matter content of Nitraria tangutorum in the artificially simulated precipitation. Acta Ecologica Sinica, 35, 4707-4715. |
| [ 任昱, 卢琦, 吴波, 李明虎 ( 2015). 不同模拟增雨下白刺比叶面积和叶干物质含量的比较. 生态学报, 35, 4707-4715.] | |
| [50] |
Roche P, Díaz-Burlinson N, Gachet S ( 2004). Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? Plant Ecology, 174, 37-48.
DOI URL |
| [51] | Shi YC, Zhao CZ, Song QH, Du J, Chen J, Wang JW ( 2015). Slope-related variations in twig and leaf traits of Robinia pseudoacacia in the northern mountains of Lanzhou. Chinese Journal of Plant Ecology, 39, 362-370. |
| [ 史元春, 赵成章, 宋清华, 杜晶, 陈静, 王继伟 ( 2015). 兰州北山刺槐枝叶性状的坡向差异性. 植物生态学报, 39, 362-370.] | |
| [52] |
Shipley B, Lechowicz MJ, Wright IJ ( 2006). Fundamental trade-offs generating the worldwide leaf economics spectrum. Ecology, 87, 535-541.
DOI URL |
| [53] | Sims DA, Pearcy RW ( 1992). Response of leaf anatomy and photosynthetic capacity in Alocasia macrorrhiza(Araceae) to a transfer from low to high light. American Journal of Botany, 79, 449-455. |
| [54] |
Smart SM, Glanville HC, Blanes MDC, Mercado LM, Emmett BA, Jones DL, Cosby BJ, Marrs RH, Butler A, Marshall MR, Reinsch S, Herrero-Jáuregui C, Hodgson JG, Field K ( 2017). Leaf dry matter content is better at predicting above-ground net primary production than specific leaf area. Functional Ecology, 31, 1336-1344.
DOI URL |
| [55] |
Sterck FJ, Poorter L, Schieving F ( 2006). Leaf traits determine the growth-survival trade-off across rain forest tree species. The American Naturalist, 167, 756-765.
DOI URL PMID |
| [56] |
Torrez V, J?rgensen PM, Zanne AE ( 2013). Specific leaf area: A predictive model using dried samples. Australian Journal of Botany, 61, 350-357.
DOI URL |
| [57] |
Wang XH, Zhang J, Zhang ZX ( 2000). Leaf longevity of evergreen broad-leaved species of Tiantong National Forest Park, Zhejiang Province. Acta Phytoecologica Sinica, 24, 625-629.
DOI URL |
|
[ 王希华, 张婕, 张正祥 ( 2000). 浙江天童国家森林公园主要常绿阔叶树种叶子寿命的研究. 植物生态学报, 24, 625-629.]
DOI URL |
|
| [58] |
Westoby M (1998). A leaf-height-seed (lhs) plant ecology strategy scheme. Plant and Soil, 199, 213-227.
DOI URL |
| [59] |
Westoby M, Falster DS, Moles AT, Vesk PA, Wright IJ ( 2002). Plant ecological strategies: Some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 33, 125-159.
DOI URL |
| [60] |
Wilson PJ, Thompson KEN, Hodgson JG ( 1999). Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 143, 155-162.
DOI URL |
| [61] |
Wirth C, Lichstein JW ( 2009). The imprint of species turnover on old-growth forest carbon balances—Insights from a trait-based model of forest dynamics. Old-Growth Forests, 81-113
DOI URL |
| [62] |
Witkowski ETF, Lamont BB ( 1991). Leaf specific mass confounds leaf density and thickness. Oecologia, 88, 486-493.
DOI URL PMID |
| [63] |
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas M-L, Niinemets Ul, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R ( 2004). The worldwide leaf economics spectrum. Nature, 428, 821-827.
DOI URL |
| [64] | Wyka T, Robakowski P, Zytkowiak R ( 2008). Leaf age as a factor in anatomical and physiological acclimative responses of Taxus baccata L. needles to contrasting irradiance environments. Photosynthesis Research, 95, 87-99. |
| [65] | Yang M, Wang SL, Zhang WD, Wang QK ( 2010). Dynamics of biomass and nutrient accumulation in a Chinese-fir plantation. Chinese Journal of Applied Ecology, 21, 1674-1680. |
| [ 杨明, 汪思龙, 张伟东, 王清奎 ( 2010). 杉木人工林生物量与养分积累动态. 应用生态学报, 21, 1674-1680.] | |
| [66] | Yu YF, Song TQ, Zeng FP, Peng WX, Wen YG, Huang CB, Wu QB, Zeng ZX, Yu Y ( 2013). Dynamics changes of biomass and its allocation in Cunninghamia lanceolata plantations of different stand ages. Chinese Journal of Ecology, 32, 1660-1666. |
| [ 俞月凤, 宋同清, 曾馥平, 彭晚霞, 温远光, 黄承标, 吴庆标, 曾昭霞, 于扬 ( 2013). 杉木人工林生物量及其分配的动态变化. 生态学杂志, 32, 1660-1666.] | |
| [67] |
Zaehle S, Friend AD ( 2010). Carbon and nitrogen cycle dynamics in the O-CN land surface model: 1. Model description, site-scale evaluation, and sensitivity to parameter estimates. Global Biogeochemical Cycles, 24, 1468-1470.
DOI URL |
| [68] |
Zhang L, Luo TX, Deng KM, Li WH ( 2008). Vertical variations in specific leaf and leaf dry matter content with canopy height in Pinus yunnanensis. Journal of Beijing Forestry University, 30(1), 40-44.
DOI URL |
|
[ 张林, 罗天祥, 邓坤枚, 李文华 ( 2008). 云南松比叶面积和叶干物质含量随冠层高度的垂直变化规律. 北京林业大学学报, 30(1), 40-44.]
DOI URL |
|
| [69] |
Zhao MF, Xiang WH, Peng CH, Tian DL ( 2009). Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model. Forest Ecology and Management, 257, 1520-1531.
DOI URL |
| [70] | Zheng YP, Zhu H, Xu HN, Xu FH, Zhou YQ ( 1991). The characteristics of growing processes of Chinese fir and division of its growth stages. Journal of Zhejiang Forestry College, 8, 219-226. |
| [ 郑勇平, 朱浩, 徐焕农, 许凤华, 周岳泉 ( 1991). 杉木生长过程特征和生长期划分. 浙江林学院学报, 8, 219-226.] | |
| [71] |
Zhou JY, He JJ, Guo ZY, Wang BB, Zhang XW, Guo CY ( 2013). A study on specific leaf area and leaf dry matter content of five dominant species in Xiangshan Mountain, Huaibei City, Anhui Province. Journal of Huaibei Normal University (Natural Science), 34(3), 51-54.
DOI URL |
|
[ 周济源, 何俊洁, 郭治远, 王贝贝, 张兴旺, 郭传友 ( 2013). 淮北相山主要优势物种比叶面积与叶干物质含量初步研究. 淮北师范大学学报(自然科学版), 34(3), 51-54.]
DOI URL |
| [1] | 吴帆, 吴晨, 张宇辉, 余恒, 魏智华, 郑蔚, 刘小飞, 陈仕东, 杨智杰, 熊德成. 增温对成熟杉木人工林不同季节细根生长、形态及生理代谢特征的影响[J]. 植物生态学报, 2023, 47(6): 856-866. |
| [2] | 周莹莹, 林华. 不同水热梯度下冠层优势树种叶片热力性状及适应策略的变化趋势[J]. 植物生态学报, 2023, 47(5): 733-744. |
| [3] | 王文伟, 韩伟鹏, 刘文文. 滨海湿地入侵植物互花米草叶片功能性状对潮位的短期响应[J]. 植物生态学报, 2023, 47(2): 216-226. |
| [4] | 谢欢, 张秋芳, 陈廷廷, 曾泉鑫, 周嘉聪, 吴玥, 林惠瑛, 刘苑苑, 尹云锋, 陈岳民. 氮添加促进丛枝菌根真菌和根系协作维持土壤磷有效性[J]. 植物生态学报, 2022, 46(7): 811-822. |
| [5] | 王广亚, 陈柄华, 黄雨晨, 金光泽, 刘志理. 着生位置对水曲柳小叶性状变异及性状间相关性的影响[J]. 植物生态学报, 2022, 46(6): 712-721. |
| [6] | 谢欢, 张秋芳, 曾泉鑫, 周嘉聪, 马亚培, 吴玥, 刘苑苑, 林惠瑛, 尹云锋, 陈岳民. 氮添加对杉木苗期磷转化和分解类真菌的影响[J]. 植物生态学报, 2022, 46(2): 220-231. |
| [7] | 张自琰, 金光泽, 刘志理. 不同区域针叶年龄对红松叶性状及相关关系的影响[J]. 植物生态学报, 2021, 45(3): 253-264. |
| [8] | 丁凯, 张毓婷, 张俊红, 柴雄, 周世水, 童再康. 不同密度杉木林对林下植被和土壤微生物群落结构的影响[J]. 植物生态学报, 2021, 45(1): 62-73. |
| [9] | 李群, 赵成章, 王继伟, 文军, 李子琴, 马俊逸. 甘肃小苏干湖盐沼湿地盐地风毛菊叶形态-光合生理特征对淹水的响应[J]. 植物生态学报, 2019, 43(8): 685-696. |
| [10] | 吕中诚, 康文星, 黄志宏, 赵仲辉, 邓湘雯. 不同林龄杉木组织迁移养分的再利用[J]. 植物生态学报, 2019, 43(5): 458-470. |
| [11] | 邹显花, 胡亚楠, 韦丹, 陈思同, 吴鹏飞, 马祥庆. 磷高效利用杉木对低磷胁迫的适应性与内源激素的相关性[J]. 植物生态学报, 2019, 43(2): 139-151. |
| [12] | 沈芳芳, 李燕燕, 刘文飞, 段洪浪, 樊后保, 胡良, 孟庆银. 长期氮沉降对杉木人工林叶、枝氮磷养分再吸收的影响[J]. 植物生态学报, 2018, 42(9): 926-937. |
| [13] | 陈日升, 康文星, 周玉泉, 田大伦, 项文化. 杉木人工林养分循环随林龄变化的特征[J]. 植物生态学报, 2018, 42(2): 173-184. |
| [14] | 陈思同, 邹显花, 蔡一冰, 韦丹, 李涛, 吴鹏飞, 马祥庆. 基于 32P示踪的不同供磷环境杉木幼苗磷的分配规律分析[J]. 植物生态学报, 2018, 42(11): 1103-1112. |
| [15] | 李群, 赵成章, 赵连春, 王建良, 张伟涛, 姚文秀. 秦王川盐沼湿地芦苇比叶面积与叶片热耗散的关联性分析[J]. 植物生态学报, 2017, 41(9): 985-994. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19