
植物生态学报 ›› 2023, Vol. 47 ›› Issue (11): 1507-1522.DOI: 10.17521/cjpe.2023.0098
所属专题: 全球变化与生态系统; 生态系统结构与功能; 生物多样性
李杰, 郝珉辉, 范春雨, 张春雨, 赵秀海( )
)
收稿日期:2023-04-10
接受日期:2023-06-15
出版日期:2023-11-20
发布日期:2023-12-22
通讯作者:
赵秀海 (作者简介:ORCID:李杰: 0009-0004-5913-7607
基金资助:
LI Jie, HAO Min-Hui, FAN Chun-Yu, ZHANG Chun-Yu, ZHAO Xiu-Hai()
Received:2023-04-10
Accepted:2023-06-15
Online:2023-11-20
Published:2023-12-22
Contact:
ZHAO Xiu-Hai(Supported by:摘要:
生物多样性对维持多种生态系统功能和提高群落抵抗扰动能力具有重要意义。选择效应和生态位互补效应是被广泛讨论的维持生态系统功能的两种机制。然而, 对于两种机制在气候变化背景下如何维持森林生态系统多功能性(EMF)的理解还不充分。该研究基于分布在中国东北寒温带和中温带天然森林的样地, 以功能性状多样性(FDq = 0)、单个和多维性状功能分散指数(FDis)代表生态位互补效应, 群落加权平均性状值(CWM)代表选择效应, 使用多元线性模型和偏最小二乘法路径模型(结构方程模型), 探究气候变化背景下不同气候区内EMF的主要影响因子和驱动机制。主要结果有: (1)在中温带森林中, 生物多样性的两种属性(树种多样性(SR)和FDq = 0)都对EMF有显著的正效应, 但FDq = 0比SR更重要。在寒温带森林中, 没有识别到生物多样性与生态系统多功能性之间的显著关系(BEMF)。(2)中温带森林群落SR对EMF的正效应被性状差异和群落加权平均最大树高(CWMHmax)介导, 选择效应和生态位互补效应两种机制同时维持EMF, 选择效应略高于互补效应。CWMHmax是影响寒温带森林EMF的主要生物因素, 选择效应是寒温带森林EMF的主要维持机制, SR对EMF的促进作用不显著, 性状差异与EMF无关。(3)由于生物多样性的“保险效应”, 中温带森林抵抗气候变化的能力更强, 气候因素对SR、性状差异、CWMHmax和EMF的影响都不显著。寒温带森林对气候变化敏感, 气候是影响EMF的重要非生物因素。更高的年平均气温和降水量显著改变了群落的性状组成(CWMHmax), 稀释了具有高竞争力和适应力性状(例如, 最大树高(Hmax))的物种对生态系统功能的贡献, 降低了选择效应。该研究结果强调了生物多样性对维持森林EMF的重要性, 证明了选择效应和生态位互补效应都是中国东北温带森林EMF的驱动机制, 并表明气候变化可能会通过改变寒温带地区森林群落性状组成(例如, CWMHmax)间接影响EMF。
李杰, 郝珉辉, 范春雨, 张春雨, 赵秀海. 东北温带森林树种和功能多样性对生态系统多功能性的影响. 植物生态学报, 2023, 47(11): 1507-1522. DOI: 10.17521/cjpe.2023.0098
LI Jie, HAO Min-Hui, FAN Chun-Yu, ZHANG Chun-Yu, ZHAO Xiu-Hai. Effect of tree species and functional diversity on ecosystem multifunctionality in temperate forests of northeast China. Chinese Journal of Plant Ecology, 2023, 47(11): 1507-1522. DOI: 10.17521/cjpe.2023.0098
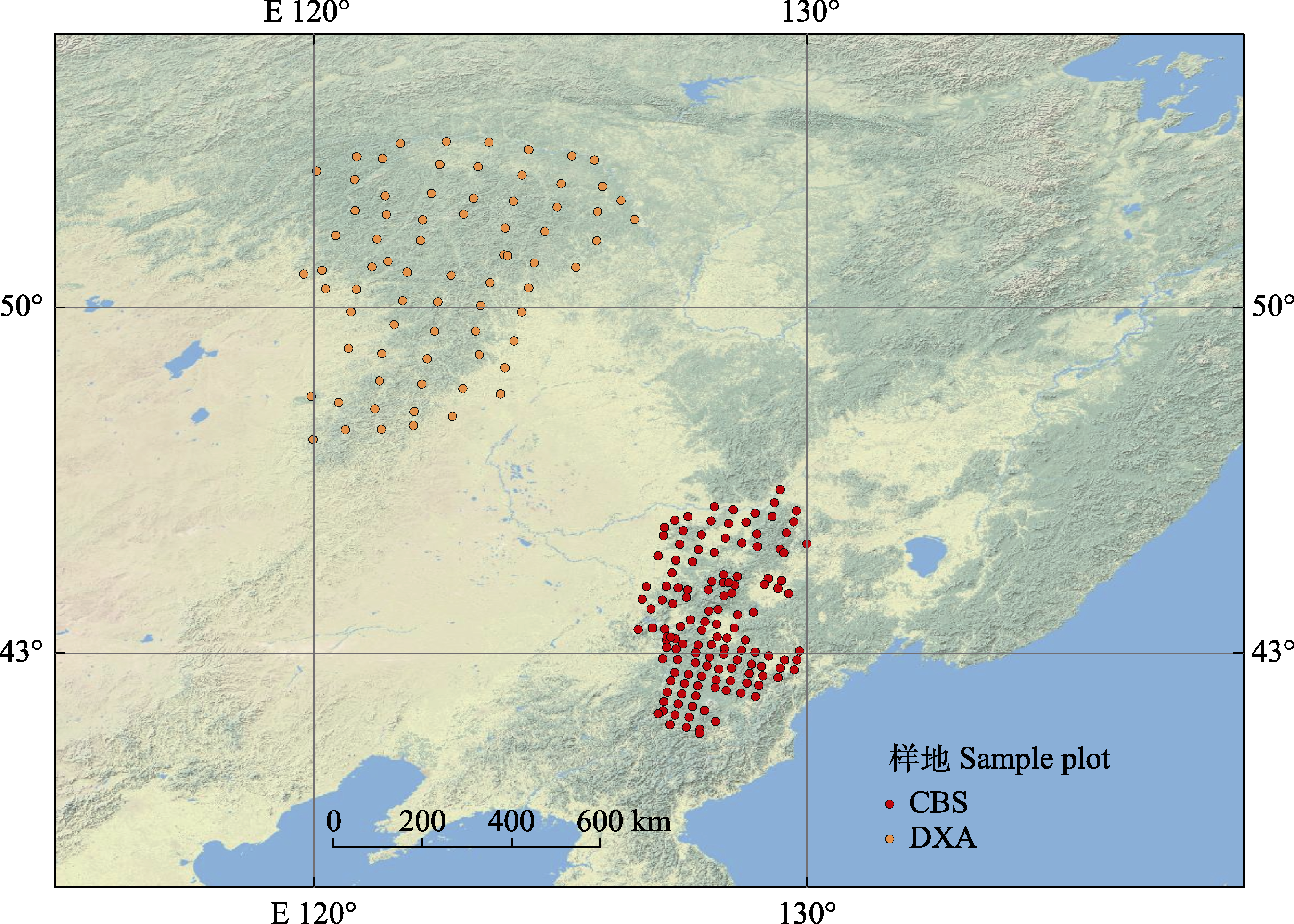
图1 东北温带森林209个森林样地的地理分布。CBS, 中温带长白山; DXA, 寒温带大兴安岭。
Fig. 1 Geographic distribution of the 209 forest plots in temperate forests of northeast China. CBS, middle temperate Changbai Mountains; DXA, cold temperate Da Hinggan Mountains.
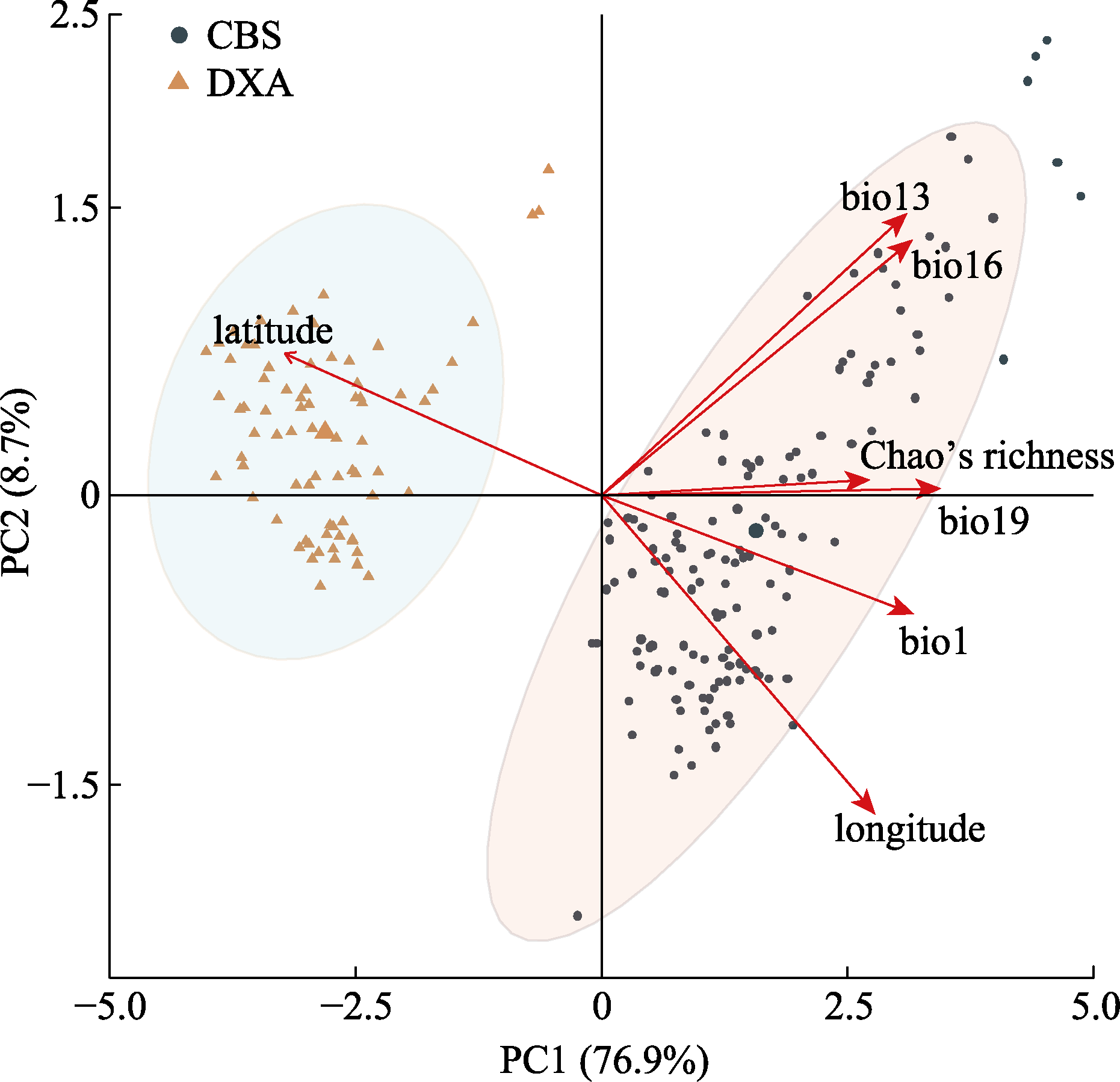
图2 东北温带森林地理位置、气候和树种多样性的主成分(PC)分析。CBS, 中温带长白山; DXA, 寒温带大兴安岭。bio1, 年平均气温; bio13, 最湿月份降水量; bio16, 最干季度降水量; bio19, 最冷季度降水量; Chao’s richness, 树种多样性; latitude, 纬度; longitude, 经度。
Fig. 2 Results of principal component (PC) analysis with geography, climate, and tree species diversity for the forest plots in temperate forests of northeast China. CBS, middle temperate Changbai Mountains; DXA, cold temperate Da Hinggan Mountains. bio1, annual mean air temperature; bio13, precipitation of the wettest month; bio16, precipitation of the driest quarter; bio19, precipitation of the coldest quarter; Chao’s richness, tree species diversity.
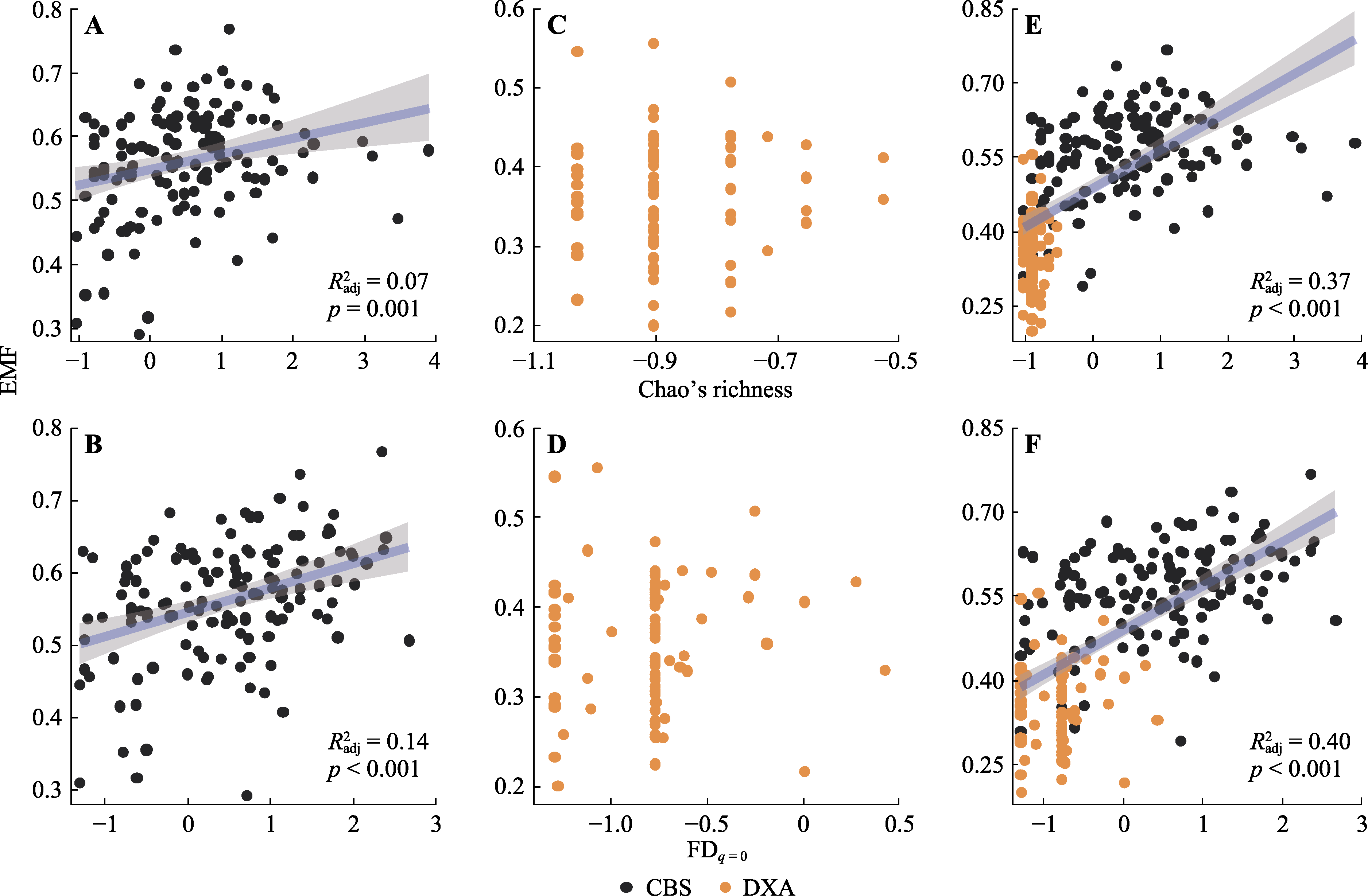
图3 东北温带森林生物多样性与生态系统多功能性(EMF)之间的关系。Chao’s richness, 树种多样性; FDq = 0, 功能性状多样性; $R_{\text{adj}}^{2}$, 模.型调整R2。灰色区域表示模型95%置信区间。CBS, 中温带长白山; DXA, 寒温带大兴安岭。
Fig. 3 Bivariate relationships between biodiversity and ecosystem multifunctionality (EMF) in temperate forests of northeast China.Chao’s richness, tree species diversity; FDq = 0, functional trait diversity;$R_{\text{adj}}^{2}$, the adjusted R2 of the model. The grey area represents the 95% confidence interval of the model. CBS, middle temperate Changbai Mountain; DXA, cold temperate Da Hinggan Mountains.
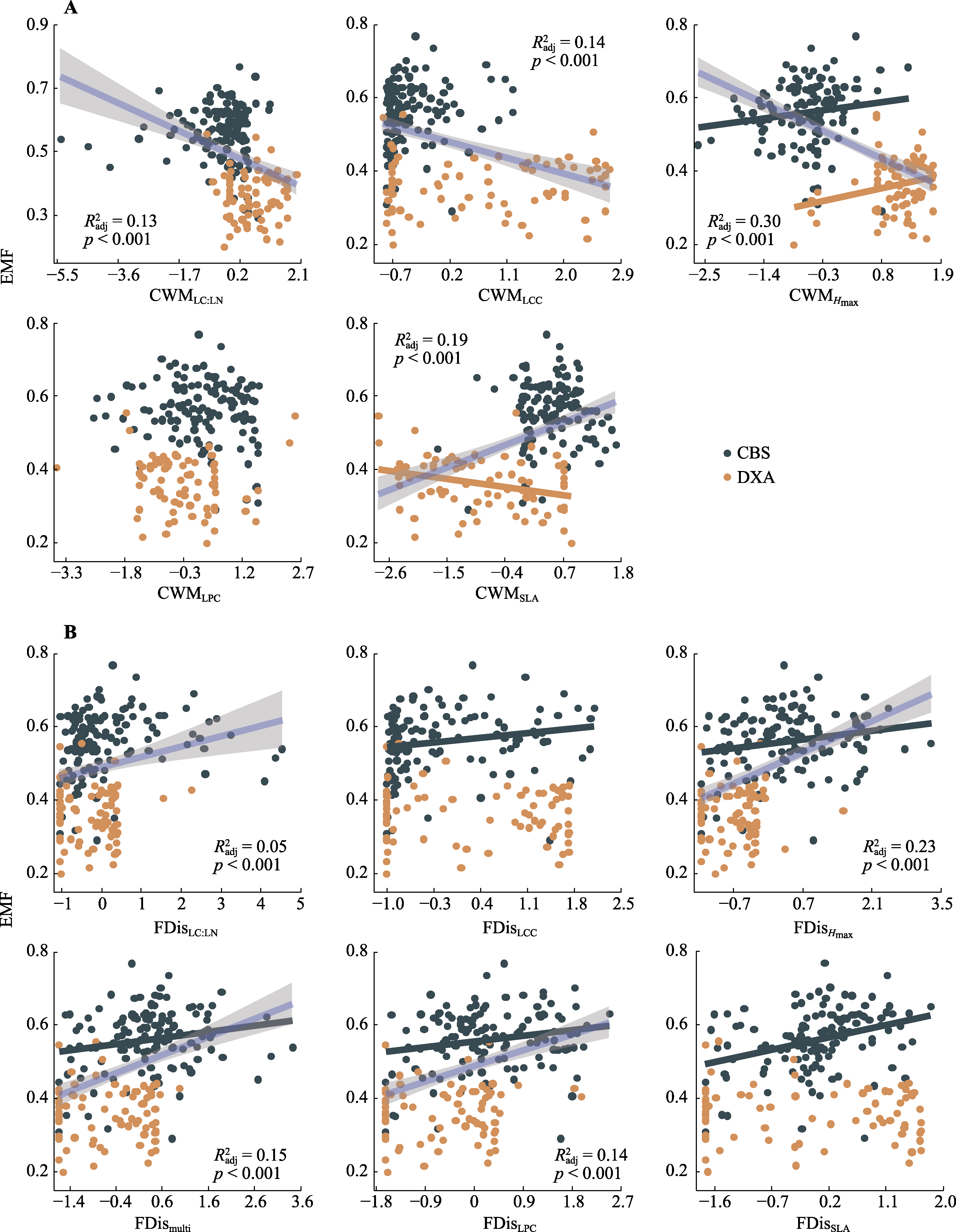
图4 东北温带森林群落功能性状变量与生态系统多功能性(EMF)之间的关系。A, 群落加权平均性状值(CWM)。B, 功能分散指数(FDis)。Hmax, 最大树高; LC:LN, 叶片碳氮比; LCC, 叶片碳含量; LPC, 叶片磷含量; multi, 多维性状功能; SLA, 比叶面积。$R_{\text{adj}}^{2}$, 模型调整R2。蓝色实线代表整个数据集上的趋势线, 灰色区域代表模型95%置信区间。CBS, 中温带长白山; DXA, 寒温带大兴安岭。
Fig. 4 Bivariate relationships between forest community trait variables and ecosystem multifunctionality (EMF) in temperate forests of northeast China. A, Community weighted mean trait values (CWM). B, Functional dispersion indices (FDis). Hmax, maximum tree height; LC:LN, leaf carbon nitrogen content ratio; LCC, leaf carbon content; LPC, leaf phosphorus content; multi, multidimensional trait; SLA, specific leaf area. R2adj, the adjusted R2 of the model. Blue solid line represents the trend line in the entire dataset, the grey area represents the 95% confidence interval of the model. CBS, middle temperate Changbai Mountains; DXA, cold temperate Da Hinggan Mountains.
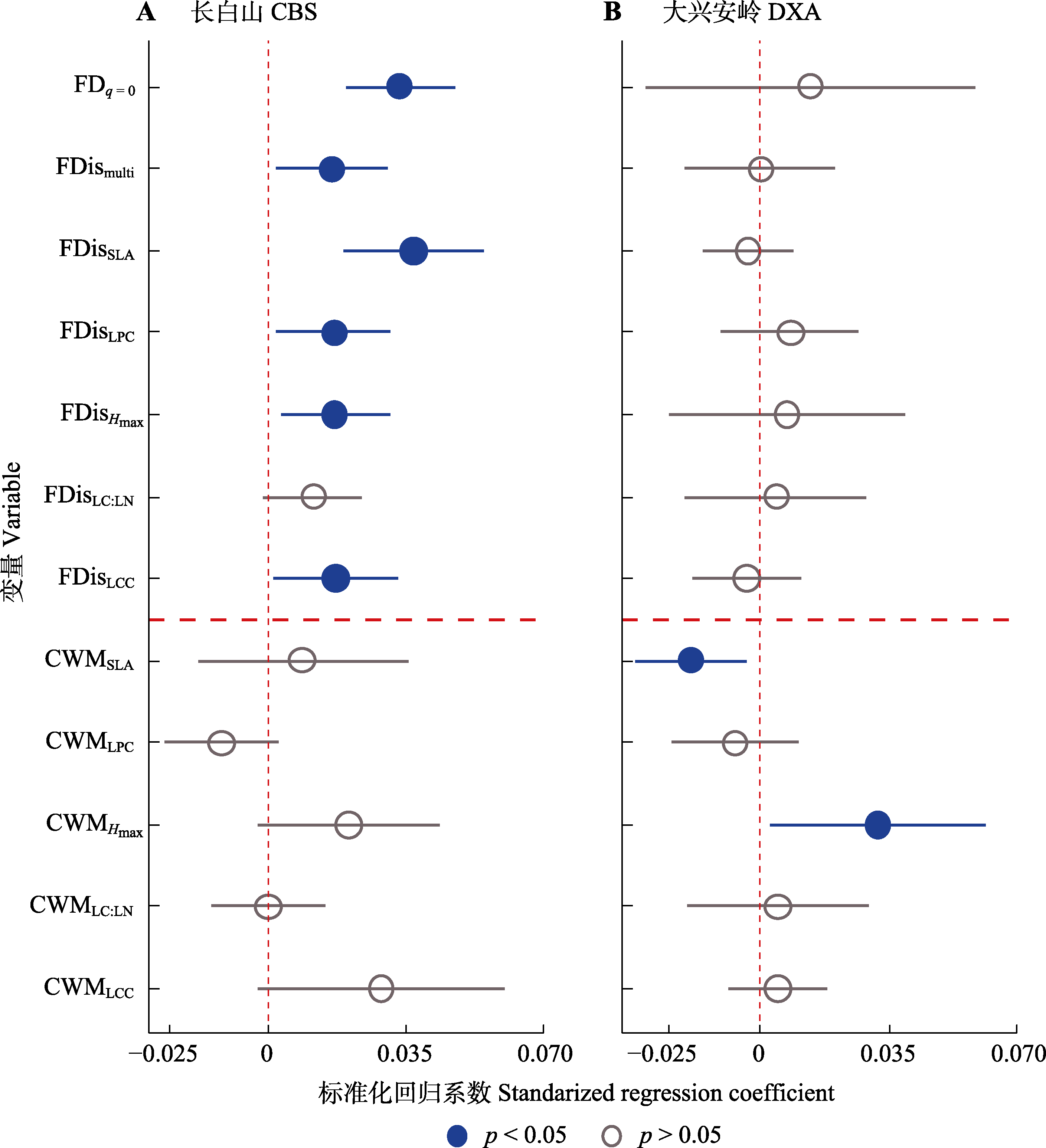
图5 东北温带森林群落功能性状变量与生态系统多功能性(EMF)之间的双变量关系。CWM, 群落加权平均性状值; FDis, 功能分散指数; FDq = 0, 功能性状多样性; Hmax, 最大树高; LCC:LNC, 叶片碳氮比; LCC, 叶片碳含量; LPC, 叶片磷含量; multi, 多维性状; SLA, 比叶面积。圆圈和短线分别代表标准化回归系数估计值和95%置信区间。
Fig. 5 Bivariate relationships between forest community trait variables and ecosystem multifunctionality (EMF) in temperate forests of northeast China. CBS, Changbai Mountains; DXA, Da Hinggan Mountains. CWM, community weighted mean trait values; FDis, functional dispersion indices; FDq = 0, functional trait diversity; Hmax, maximum tree height; LC:LN, leaf carbon nitrogen content ratio; LCC, leaf carbon content; LPC, leaf phosphorus content; multi, multidimensional trait; SLA, specific leaf area. The dots and dashes represent the estimated values of standardized regression coefficients and the 95% confidence interval, respectively.
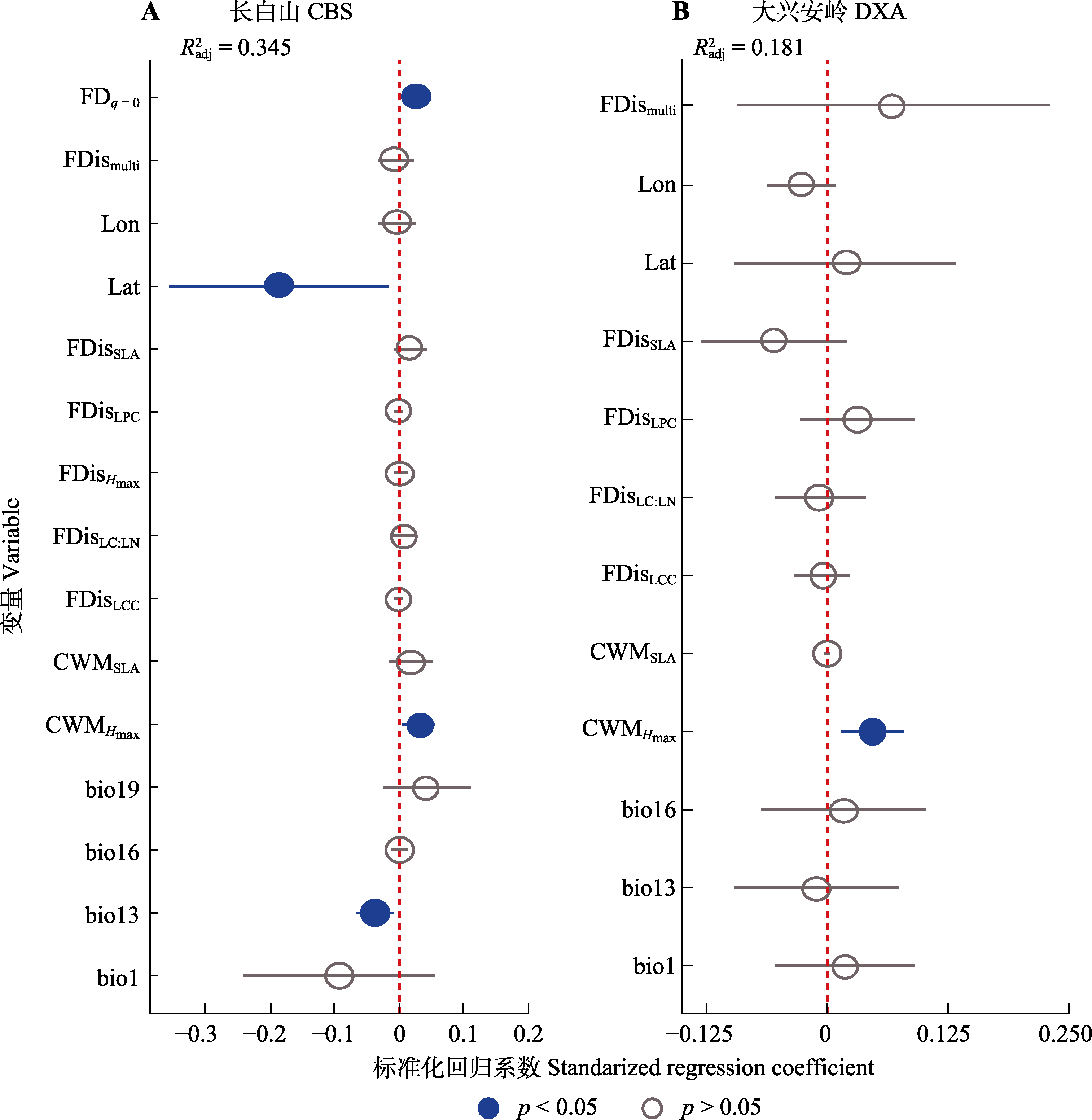
图6 东北温带森林生态系统多功能性(EMF)的最佳预测变量。bio1, 年平均气温; bio13, 最湿月份降水量; bio16, 最干季度降水量; bio19, 最冷季度降水量; CWMHmax, 群落加权平均最大树高; CWMSLA, 群落加权平均比叶面积; FDisHmax, 最大树高功能分散指数; FDisLC:LN, 叶片碳氮比功能分散指数; FDisSLA, 比叶面积功能分散指数; FDisLCC, 叶片碳含量功能分散指数; FDisLPC, 叶片磷含量功能分散指数; FDismulti, 多维性状功能分散指数; FDq = 0, 功能性状多样性; Lat, 纬度; Lon, 经度; R2adj, 模型调整R2。圆点和短线分别代表标准化回归系数估计值和95%置信区间。
Fig. 6 Optimal predictors of ecosystem multifunctionality (EMF) in temperate forests of northeast China. CBS, Changbai Mountains. DXA, Da Hinggan Mountains. bio1, annual mean air temperature; bio13, precipitation of the wettest month; bio16, precipitation of the driest quarter; bio19, precipitation of the coldest quarter; CWMHmax, maximum tree height community weighted mean trait value; CWMSLA, specific leaf area community weighted mean trait value; FDisHmax, maximum tree height functional dispersion index; FDisLC:LN, leaf carbon nitrogen content ratio functional dispersion index; FDisSLA, specific leaf area functional dispersion index; FDisLCC, leaf carbon content functional dispersion index; FDisLPC, leaf phosphorus content functional dispersion index; FDismulti, multidimensional trait functional dispersion index; FDq = 0, functional trait diversity; Lat, latitude; Lon, longitude. R2adj, the adjusted R2 of the model. The dots and dashes represent the estimated values of standardized regression coefficients and the 95% confidence interval, respectively.
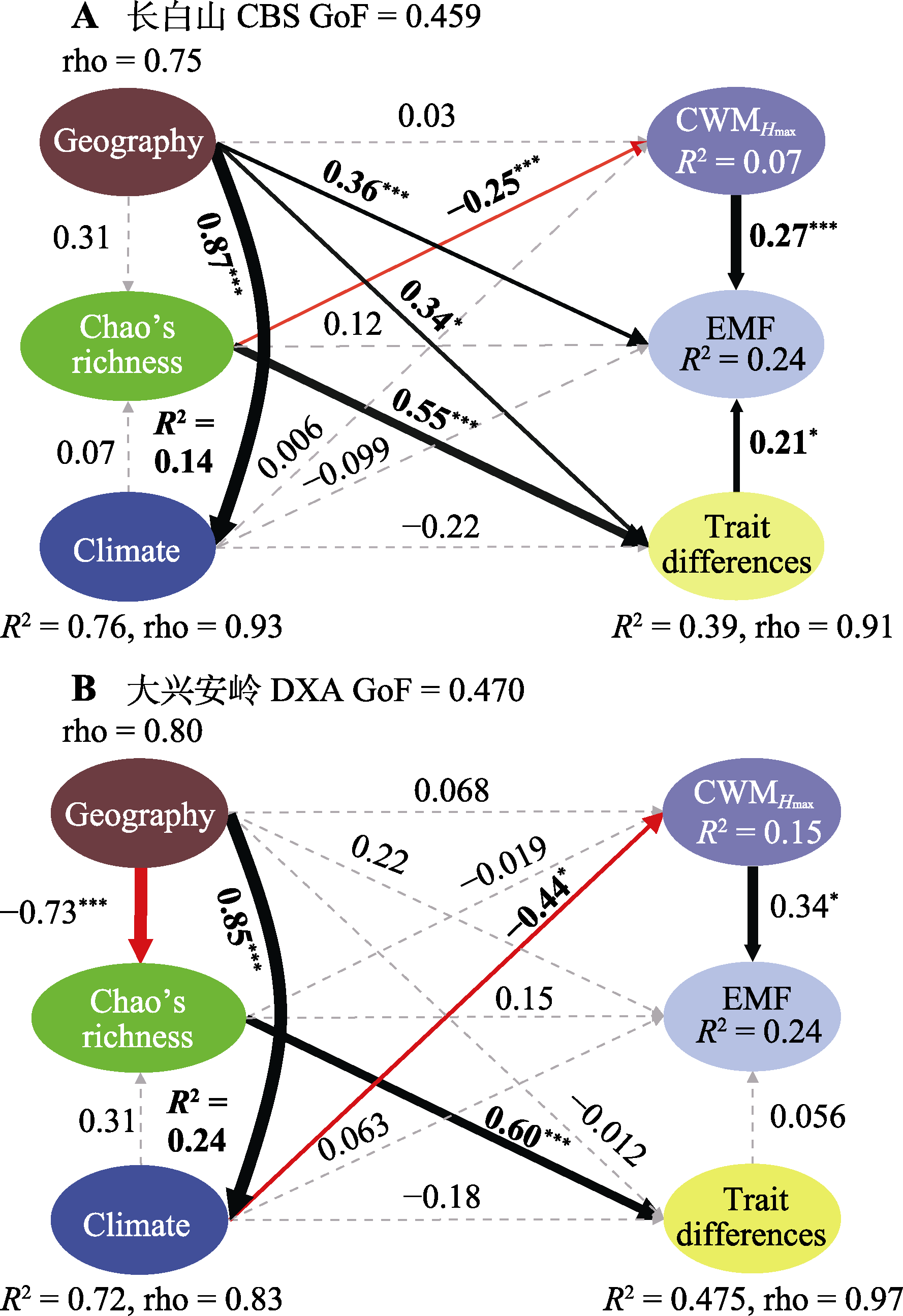
图7 东北温带森林生物和非生物因素对生态系统多功能性(EMF)影响的结构方程模型图。Chao’s richness, 树种多样性; Climate, 气候因素; CWMHmax, 群落加权平均最大树高; Geography, 地理因素; GoF, 模型拟合优度; rho, Dillon-Goldstein’s rho值; Trait differences, 性状差异。黑色、红色实线和虚线表示路径正、负和不显著。箭头线附近的数值表示标准化路径系数(β), 并给出因变量R2。*, p < 0.05; ***, p < 0.001。
Fig. 7 Structural equation model about the impact of biotic and abiotic factors on ecosystem multifunctionality (EMF) in northeast temperate forests. CBS, Changbai Mountains. DXA, Da Hinggan Mountains. Chao’s richness, tree species diversity; CWMHmax, community weighted mean maximum tree height value; GoF, model goodness-of-fit value; rho, Dillon-Goldstein’s rho value. Solid black, red, and dashed lines represent positive, negative, and non-significant paths, respectively. Standardized path coefficients (β) are indicated near the arrow lines, and response variables’ R2 are provided. *, p < 0.05; ***, p < 0.001.
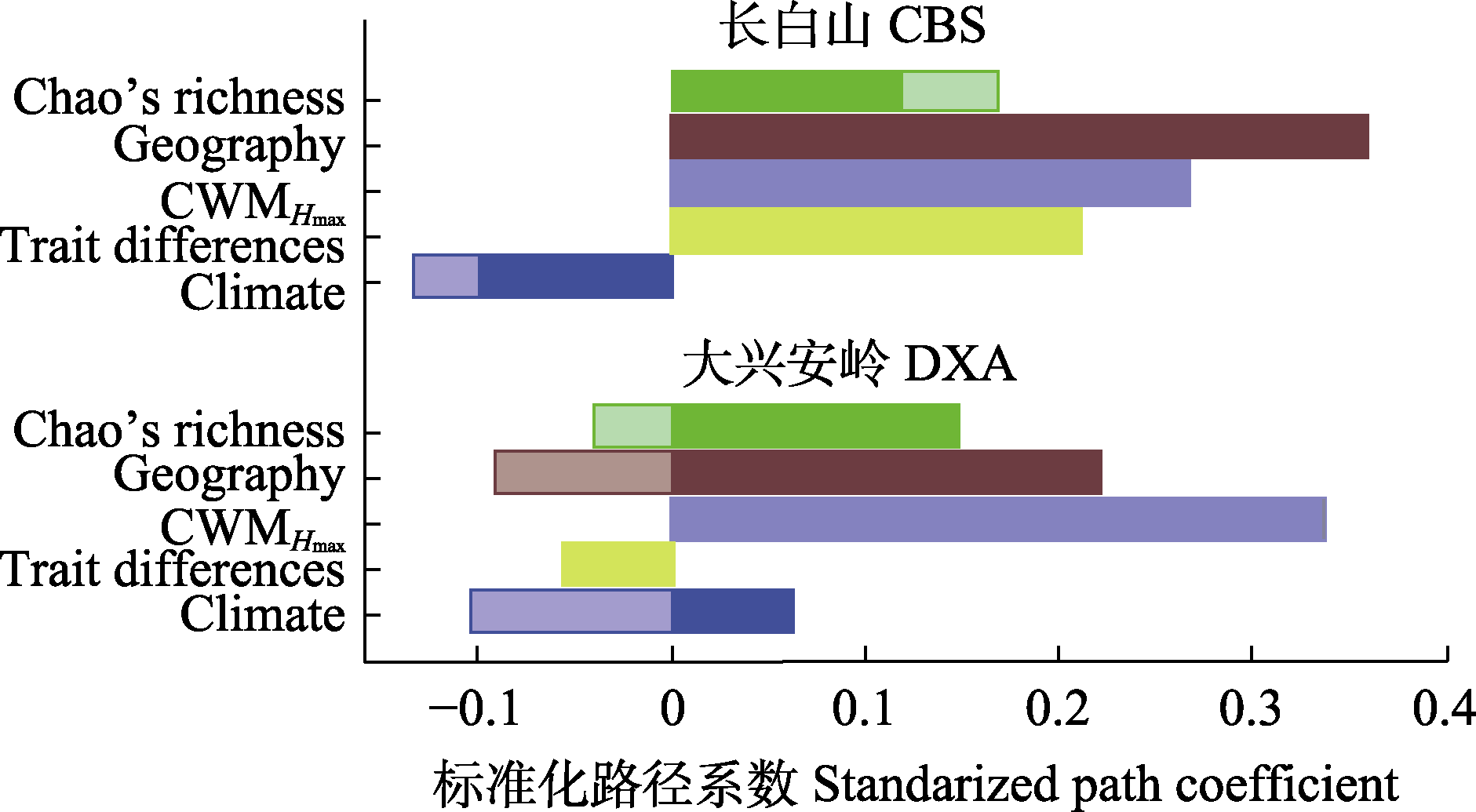
图8 东北温带森林生物和非生物因素对生态系统多功能性(EMF)的直接和间接效应。Chao’s richness, 树种多样性; Climate, 气候因素(潜在变量); CWMHmax, 群落加权平均最大树高; Geography, 地理因素(潜在变量); Trait differences, 性状差异(潜在变量)。深色区域表示直接效应, 透明区域表示间接效应。
Fig. 8 Direct and indirect effects of biotic and abiotic factors on ecosystem multifunctionality (EMF) in temperate forests of northeast China. CBS, the Changbai Mountain; DXA, the Da Hinggan Mountains. Chao’s richness, tree species diversity; Climate, climate factor (latent variable); CWMHmax, community weighted mean maximum tree height value; Geography, geography factor (latent variable); Trait differences, trait differences (latent variable). The dark and transparent bars represent direct and indirect effects, respectively.
| [1] |
Byrnes JEK, Gamfeldt L, Isbell F, Lefcheck JS, Griffin JN, Hector A, Cardinale BJ, Hooper DU, Dee LE, Emmett Duffy J (2014). Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods in Ecology and Evolution, 5, 111-124.
DOI URL |
| [2] |
Cadotte MW (2017). Functional traits explain ecosystem function through opposing mechanisms. Ecology Letters, 20, 989-996.
DOI PMID |
| [3] | Chao A, Chiu CH, Villéger S, Sun IF, Thorn S, Lin YC, Chiang JM, Sherwin WB (2019). An attribute-diversity approach to functional diversity, functional beta diversity, and related (dis)similarity measures. Ecological Monographs, 89, e01343. DOI: 10.1002/ecm.1343. |
| [4] |
Chao A, Henderson PA, Chiu CH, Moyes F, Hu KH, Dornelas M, Magurran AE (2021). Measuring temporal change in alpha diversity: a framework integrating taxonomic, phylogenetic and functional diversity and the iNEXT.3D standardization. Methods in Ecology and Evolution, 12, 1926-1940.
DOI URL |
| [5] |
Crous KY, O’Sullivan OS, Zaragoza-Castells J, Bloomfield KJ, Negrini ACA, Meir P, Turnbull MH, Griffin KL, Atkin OK (2017). Nitrogen and phosphorus availabilities interact to modulate leaf trait scaling relationships across six plant functional types in a controlled-environment study. New Phytologist, 215, 992-1008.
DOI PMID |
| [6] |
Da RH, Fan CY, Zhang CY, Zhao XH, von Gadow K (2023). Are absorptive root traits good predictors of ecosystem functioning? A test in a natural temperate forest. New Phytologist, 239, 75-86.
DOI URL |
| [7] |
Fang JY, Guo ZD, Hu HF, Kato T, Muraoka H, Son Y (2014). Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth. Global Change Biology, 20, 2019-2030.
DOI PMID |
| [8] |
Fick SE, Hijmans RJ (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37, 4302-4315.
DOI URL |
| [9] |
Fox JW (2005). Interpreting the “selection effect” of biodiversity on ecosystem function. Ecology Letters, 8, 846-856.
DOI URL |
| [10] |
Gamfeldt L, Hillebrand H, Jonsson PR (2008). Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology, 89, 1223-1231.
DOI PMID |
| [11] | Gamfeldt L, Roger F (2017). Revisiting the biodiversity- ecosystem multifunctionality relationship. Nature Ecology & Evolution, 1, 168. DOI: 10.1038/s41559-017-0168. |
| [12] | Gamfeldt L, Snäll T, Bagchi R, Jonsson M, Gustafsson L, Kjellander P, Ruiz-Jaen MC, Fröberg M, Stendahl J, Philipson CD, Mikusiński G, Andersson E, Westerlund B, Andrén H, Moberg F, et al. (2013). Higher levels of multiple ecosystem services are found in forests with more tree species. Nature Communications, 4, 1340. DOI: 10.1038/ncomms2328. |
| [13] |
Godlee JL, Ryan CM, Bauman D, Bowers SJ, Carreiras JMB, Chisingui AV, Cromsigt JPGM, Druce DJ, Finckh M, Gonçalves FM, Holdo RM, Makungwa S, McNicol IM, Mitchard ETA, Muchawona A, et al. (2021). Structural diversity and tree density drive variation in the biodiversity-ecosystem function relationship of woodlands and savannas. New Phytologist, 232, 579-594.
DOI PMID |
| [14] |
Grman E, Zirbel CR, Bassett T, Brudvig LA (2018). Ecosystem multifunctionality increases with beta diversity in restored prairies. Oecologia, 188, 837-848.
DOI PMID |
| [15] | Gross N, Le Bagousse-Pinguet Y, Liancourt P, Berdugo M, Gotelli NJ, Maestre FT (2017). Functional trait diversity maximizes ecosystem multifunctionality. Nature Ecology & Evolution, 1, 132. DOI: 10.1038/s41559-017-0132. |
| [16] |
Guo C, Yan ER, Cornelissen JHC (2022). Size matters for linking traits to ecosystem multifunctionality. Trends in Ecology & Evolution, 37, 803-813.
DOI URL |
| [17] |
Hagan JG, Vanschoenwinkel B, Gamfeldt L (2021). We should not necessarily expect positive relationships between biodiversity and ecosystem functioning in observational field data. Ecology Letters, 24, 2537-2548.
DOI URL |
| [18] |
Hao MH, Messier C, Geng Y, Zhang CY, Zhao XH, von Gadow K (2020). Functional traits influence biomass and productivity through multiple mechanisms in a temperate secondary forest. European Journal of Forest Research, 139, 959-968.
DOI |
| [19] | Hautier Y, Isbell F, Borer ET, Seabloom EW, Stanley H W, Lind EM, MacDougall AS, Stevens CJ, Adler PB, Alberti J, Bakker JD, Brudvig LA, Buckley YM, Cadotte M, Caldeira MC, et al. (2018). Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality. Nature Ecology & Evolution, 2, 50-56. |
| [20] | Hengl T, Mendes de Jesus J, Heuvelink GBM, Ruiperez Gonzalez M, Kilibarda M, Blagotić A, Shangguan W, Wright MN, Geng XY, Bauer-Marschallinger B, Guevara MA, Vargas R, MacMillan RA, Batjes NH, Leenaars JGB, et al. (2017). SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE, 12, e0169748. DOI: 10.1371/journal.pone.0169748. |
| [21] | Hu WG, Ran JZ, Dong LW, Du QJ, Ji MF, Yao SR, Sun Y, Gong CM, Hou QQ, Gong HY, Chen RF, Lu JL, Xie SB, Wang ZQ, Huang H, et al. (2021). Aridity-driven shift in biodiversity-soil multifunctionality relationships. Nature Communications, 12, 5350. DOI: 10.1038/s41467-021- 25641-0. |
| [22] | Hu Y, Li XY, Guo AX, Yue P, Guo XX, Lv P, Zhao SL, Zuo XA (2022). Species diversity is a strong predictor of ecosystem multifunctionality under altered precipitation in desert steppes. Ecological Indicators, 137, 108762. DOI: 10.1016/j.ecolind.2022.108762. |
| [23] | Huang XB, Su JR, Li SF, Liu WD, Lang XD (2019). Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Scientific Reports, 9, 6979. DOI: 10.1038/s41598-019-43475-1. |
| [24] |
Huang YY, Chen YX, Castro-Izaguirre N, Baruffol M, Brezzi M, Lang A, Li Y, Härdtle W, von Oheimb G, Yang XF, Liu XJ, Pei KQ, Both S, Yang B, Eichenberg D, et al. (2018). Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science, 362, 80-83.
DOI PMID |
| [25] |
Isbell F, Craven D, Connolly J, Loreau M, Schmid B, Beierkuhnlein C, Bezemer TM, Bonin C, Bruelheide H, de Luca E, Ebeling A, Griffin JN, Guo QF, Hautier Y, Hector A, et al. (2015). Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature, 526, 574-577.
DOI |
| [26] |
Isbell FI, Polley HW, Wilsey BJ (2009). Biodiversity, productivity and the temporal stability of productivity: patterns and processes. Ecology Letters, 12, 443-451.
DOI PMID |
| [27] | Jing X, Sanders NJ, Shi Y, Chu HY, Classen AT, Zhao K, Chen LT, Shi Y, Jiang YX, He JS (2015). The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nature Communications, 6, 8159. DOI: 10.1038/ncomms9159. |
| [28] | Kattge J, Díaz S, Lavorel S, Prentice IC, Leadley P, Bönisch G, Garnier E, Westoby M, Reich PB, Wright IJ, Cornelissen JHC, Violle C, Harrison SP, van Bodegom PM, Reichstein M, et al. (2011). TRY—A global database of plant traits. Global Change Biology, 17, 2905-2935. |
| [29] |
Kraft NJB, Godoy O, Levine JM (2015). Plant functional traits and the multidimensional nature of species coexistence. Proceedings of the National Academy of Sciences of the United States of America, 112, 797-802.
DOI PMID |
| [30] |
Laliberté E, Legendre P (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology, 91, 299-305.
PMID |
| [31] |
Lasky JR, Uriarte M, Boukili VK, Erickson DL, Kress WJ, Chazdon RL (2014). The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession. Ecology Letters, 17, 1158-1167.
DOI PMID |
| [32] |
Li JL, Chen XP, Niklas KJ, Sun J, Wang ZY, Zhong QL, Hu DD, Cheng DL (2022). A whole-plant economics spectrum including bark functional traits for 59 subtropical woody plant species. Journal of Ecology, 110, 248-261.
DOI URL |
| [33] | Li SF, Huang XB, Lang XD, Shen JY, Xu FD, Su JR (2020). Cumulative effects of multiple biodiversity attributes and abiotic factors on ecosystem multifunctionality in the Jinsha River valley of southwestern China. Forest Ecology and Management, 472, 118281. DOI: 10.1016/j.foreco.2020.118281. |
| [34] | Li SF, Liu WD, Lang XD, Huang XB, Su JR (2021). Species richness, not abundance, drives ecosystem multifunctionality in a subtropical coniferous forest. Ecological Indicators, 120, 106911. DOI: 10.1016/j.ecolind.2020.106911. |
| [35] |
Loreau M, de Mazancourt C (2013). Biodiversity and ecosystem stability: a synthesis of underlying mechanisms. Ecology Letters, 16, 106-115.
DOI URL |
| [36] |
Loreau M, Hector A (2001). Partitioning selection and complementarity in biodiversity experiments. Nature, 412, 72-76.
DOI URL |
| [37] |
Mensah S, du Toit B, Seifert T (2018). Diversity-biomass relationship across forest layers: implications for niche complementarity and selection effects. Oecologia, 187, 783-795.
DOI PMID |
| [38] | Moi DA, Antoniazi Evangelista HB, Mormul RP, Evangelista LR, Thomaz SM (2021). Ecosystem multifunctionality and stability are enhanced by macrophyte richness in mesocosms. Aquatic Sciences, 83, 53. DOI: 10.1007/s00027-021-00808-5. |
| [39] | Moi DA, Lansac-Tôha FM, Romero GQ, Sobral-Souza T, Cardinale BJ, Kratina P, Perkins DM, Teixeira de Mello F, Jeppesen E, Heino J, Lansac-Tôha FA, Velho LFM, Mormul RP (2022). Human pressure drives biodiversity- multifunctionality relationships in large Neotropical wetlands. Nature Ecology & Evolution, 6, 1279-1289. |
| [40] |
Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte MS, Cornwell WK, Craine JM, Gurvich DE, Urcelay C, Veneklaas EJ, Reich PB, Poorter L, Wright IJ, et al. (2013). New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 61, 167-234.
DOI URL |
| [41] |
Qiao XT, Lamy T, Wang SP, Hautier Y, Geng Y, White HJ, Zhang NL, Zhang ZH, Zhang CY, Zhao XH, von Gadow K (2023). Latitudinal patterns of forest ecosystem stability across spatial scales as affected by biodiversity and environmental heterogeneity. Global Change Biology, 29, 2242-2255.
DOI URL |
| [42] |
Ratcliffe S, Liebergesell M, Ruiz-Benito P, Madrigal González J, Muñoz Castañeda JM, Kändler G, Lehtonen A, Dahlgren J, Kattge J, Peñuelas J, Zavala MA, Wirth C (2016). Modes of functional biodiversity control on tree productivity across the European continent. Global Ecology and Biogeography, 25, 251-262.
DOI URL |
| [43] | Roscher C, Schumacher J, Gubsch M, Lipowsky A, Weigelt A, Buchmann N, Schmid B, Schulze ED (2012). Using plant functional traits to explain diversity-productivity relationships. PLoS ONE, 7, e36760. DOI: 10.1371/journal.pone.0036760. |
| [44] |
Scherer-Lorenzen M, Palmborg C, Prinz A, Schulze ED (2003). The role of plant diversity and composition for nitrate leaching in grasslands. Ecology, 84, 1539-1552.
DOI URL |
| [45] |
Shi XZ, Wang JQ, Lucas-Borja ME, Wang ZY, Li X, Huang ZQ (2021). Microbial diversity regulates ecosystem multifunctionality during natural secondary succession. Journal of Applied Ecology, 58, 2833-2842.
DOI URL |
| [46] |
Stone R (2006). A threatened nature reserve breaks down Asian borders. Science, 313, 1379-1380.
DOI URL |
| [47] | Tenenhaus M, Vinzi VE, Chatelin YM, Lauro C (2005). PLS path modeling. Computational Statistics & Data Analysis, 48, 159-205. |
| [48] |
Tobner CM, Paquette A, Gravel D, Reich PB, Williams LJ, Messier C (2016). Functional identity is the main driver of diversity effects in young tree communities. Ecology Letters, 19, 638-647.
DOI PMID |
| [49] |
Valencia E, Maestre FT, Le Bagousse-Pinguet Y, Quero JL, Tamme R, Börger L, García-Gómez M, Gross N (2015). Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands. New Phytologist, 206, 660-671.
DOI PMID |
| [50] |
van der Plas F (2019). Biodiversity and ecosystem functioning in naturally assembled communities. Biological Reviews, 94, 1220-1245.
DOI URL |
| [51] | van der Plas F, Manning P, Allan E, Scherer-Lorenzen M, Verheyen K, Wirth C, Zavala MA, Hector A, Ampoorter E, Baeten L, Barbaro L, Bauhus J, Benavides R, Benneter A, Berthold F, et al. (2016a). Jack-of-all-trades effects drive biodiversity-ecosystem multifunctionality relationships in European forests. Nature Communications, 7, 11109. DOI: 10.1038/ncomms11109. |
| [52] | van der Plas F, Manning P, Soliveres S, Allan E, Scherer-Lorenzen M, Verheyen K, Wirth C, Zavala MA, Ampoorter E, Baeten L, Barbaro L, Bauhus J, Benavides R, Benneter A, Bonal D, et al. (2016b). Biotic homogenization can decrease landscape-scale forest multifunctionality. Proceedings of the National Academy of Sciences of the United States of America, 113, 3557-3562. |
| [53] |
van der Plas F, Ratcliffe S, Ruiz-Benito P, Scherer-Lorenzen M, Verheyen K, Wirth C, Zavala MA, Ampoorter E, Baeten L, Barbaro L, Bastias CC, Bauhus J, Benavides R, Benneter A, Bonal D, et al. (2018). Continental mapping of forest ecosystem functions reveals a high but unrealised potential for forest multifunctionality. Ecology Letters, 21, 31-42.
DOI PMID |
| [54] | Wang WY, Hou YT, Pan WH, Vinay N, Mo F, Liao YC, Wen XX (2021). Continuous application of conservation tillage affects in situ N2O emissions and nitrogen cycling gene abundances following nitrogen fertilization. Soil Biology & Biochemistry, 157, 108239. DOI: 10.1016/j.soilbio.2021.108239. |
| [55] | Wondimu MT, Nigussie ZA, Yusuf MM (2021). Tree species diversity predicts aboveground carbon storage through functional diversity and functional dominance in the dry evergreen Afromontane forest of Hararghe highland, Southeast Ethiopia. Ecological Processes, 10, 47. DOI: 10.1186/s13717-021-00322-4. |
| [56] | Wu ZF, Zhang ZH, Wang J (2019). Estimating the productive potential of five natural forest types in northeastern China. Forest Ecosystems, 6, 42. DOI: 10.1186/s40663-019-0204-0. |
| [57] |
Yachi S, Loreau M (1999). Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America, 96, 1463-1468.
DOI PMID |
| [58] | Yan YZ, Zhang Q, Buyantuev A, Liu QF, Niu JM (2020). Plant functional β diversity is an important mediator of effects of aridity on soil multifunctionality. Science of the Total Environment, 726, 138529. DOI: 10.1016/j.scitotenv.2020.138529. |
| [59] |
Yuan ZQ, Ali A, Loreau M, Ding F, Liu SF, Sanaei A, Zhou WM, Ye J, Lin F, Fang S, Hao ZQ, Wang XG, Le Bagousse-Pinguet Y (2021). Divergent above- and below-ground biodiversity pathways mediate disturbance impacts on temperate forest multifunctionality. Global Change Biology, 27, 2883-2894.
DOI PMID |
| [60] |
Yuan ZQ, Ali A, Ruiz-Benito P, Jucker T, Mori AS, Wang SP, Zhang XK, Li H, Hao ZQ, Wang XG, Loreau M (2020). Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. Journal of Ecology, 108, 2012-2024.
DOI URL |
| [61] | Yue QM, Hao MH, Geng Y, Wang XR, von Gadow K, Zhang CY, Zhao XH, Gao LS (2022). Evaluating alternative hypotheses behind biodiversity and multifunctionality relationships in the forests of Northeastern China. Forest Ecosystems, 9, 100027. DOI: 10.1016/j.fecs.2022.100027. |
| [62] |
Zhang CY, He FL, Zhang ZH, Zhao XH, von Gadow K (2020). Latitudinal gradients and ecological drivers of β-diversity vary across spatial scales in a temperate forest region. Global Ecology and Biogeography, 29, 1257-1264.
DOI URL |
| [63] |
Zirbel CR, Bassett T, Grman E, Brudvig LA (2017). Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration. Journal of Applied Ecology, 54, 1070-1079.
DOI URL |
| [1] | 陈以恒 玉素甫江·如素力 阿卜杜热合曼·吾斯曼. 2001-2020年天山新疆段草地植被覆盖度时空变化及驱动因素分析[J]. 植物生态学报, 2024, 48(5): 561-576. |
| [2] | 张计深, 史新杰, 刘宇诺, 吴阳, 彭守璋. 气候变化下中国潜在自然植被生态系统碳储量动态[J]. 植物生态学报, 2024, 48(4): 428-444. |
| [3] | 臧妙涵, 王传宽, 梁逸娴, 刘逸潇, 上官虹玉, 全先奎. 基于纬度移栽的落叶松叶、枝、根生态化学计量特征对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 469-482. |
| [4] | 梁逸娴, 王传宽, 臧妙涵, 上官虹玉, 刘逸潇, 全先奎. 落叶松径向生长和生物量分配对气候变暖的响应[J]. 植物生态学报, 2024, 48(4): 459-468. |
| [5] | 吴茹茹, 刘美珍, 谷仙, 常馨月, 郭立月, 蒋高明, 祁如意. 气候变化对巨柏适宜生境分布的潜在影响和预测[J]. 植物生态学报, 2024, 48(4): 445-458. |
| [6] | 杨宇萌, 来全, 刘心怡. 气候变化和人类活动对内蒙古植被总初级生产力的定量影响[J]. 植物生态学报, 2024, 48(3): 306-316. |
| [7] | 张启, 程雪寒, 王树芝. 北京西山老龄树记载的森林干扰历史[J]. 植物生态学报, 2024, 48(3): 341-348. |
| [8] | 吕自立, 刘彬, 常凤, 马紫荆, 曹秋梅. 巴音布鲁克高寒草甸植物功能多样性与生态系统多功能性关系沿海拔梯度的变化[J]. 植物生态学报, 2023, 47(6): 822-832. |
| [9] | 任培鑫, 李鹏, 彭长辉, 周晓路, 杨铭霞. 洞庭湖流域植被光合物候的时空变化及其对气候变化的响应[J]. 植物生态学报, 2023, 47(3): 319-330. |
| [10] | 魏瑶, 马志远, 周佳颖, 张振华. 模拟增温改变青藏高原植物繁殖物候及植株高度[J]. 植物生态学报, 2022, 46(9): 995-1004. |
| [11] | 党宏忠, 张学利, 韩辉, 石长春, 葛玉祥, 马全林, 陈帅, 刘春颖. 樟子松固沙林林水关系研究进展及对营林实践的指导[J]. 植物生态学报, 2022, 46(9): 971-983. |
| [12] | 李肖, PIALUANG Bounthong, 康文辉, 冀晓东, 张海江, 薛治国, 张志强. 近几十年来冀西北山地白桦次生林径向生长对气候变化的响应[J]. 植物生态学报, 2022, 46(8): 919-931. |
| [13] | 苏启陶, 杜志喧, 周兵, 廖永辉, 王呈呈, 肖宜安. 牯岭凤仙花及其传粉昆虫在中国的潜在分布区域分析[J]. 植物生态学报, 2022, 46(7): 785-796. |
| [14] | 胡潇飞, 魏临风, 程琦, 吴星麒, 倪健. 青藏高原地区气候图解数据集[J]. 植物生态学报, 2022, 46(4): 484-492. |
| [15] | 原媛, 母艳梅, 邓钰洁, 李鑫豪, 姜晓燕, 高圣杰, 查天山, 贾昕. 植被覆盖度和物候变化对典型黑沙蒿灌丛生态系统总初级生产力的影响[J]. 植物生态学报, 2022, 46(2): 162-175. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2026 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19
![]()