
植物生态学报 ›› 2018, Vol. 42 ›› Issue (11): 1082-1093.DOI: 10.17521/cjpe.2018.0082
所属专题: 青藏高原植物生态学:种群生态学
周天阳1,2,3,NARAYAN Prasad Gaire4,8,廖礼彬1,2,郑莉莉2,5,王金牛1,3,6,*( ),孙建5,魏彦强7,谢雨1,吴彦1,3,*()
),孙建5,魏彦强7,谢雨1,吴彦1,3,*()
收稿日期:2018-04-11
接受日期:2018-11-04
出版日期:2018-11-20
发布日期:2019-03-13
通讯作者:
王金牛,吴彦
基金资助:
ZHOU Tian-Yang1,2,3,NARAYAN Prasad Gaire4,8,LIAO Li-Bin1,2,ZHENG Li-Li2,5,WANG Jin-Niu1,3,6,*(),SUN Jian5,WEI Yan-Qiang7,XIE Yu1,WU Yan1,3,*()
Received:2018-04-11
Accepted:2018-11-04
Online:2018-11-20
Published:2019-03-13
Contact:
Jin-Niu WANG,Yan WU
Supported by:摘要:
热量匮乏是高山树线的主要成因, 在全球变暖趋势下对高山树线及其建群种的生态学过程及特征的研究具有重要意义。该文以青藏高原东缘的折多山和剪子弯山两处高山树线(海拔分别为4 265 m和4 425 m)作为研究对象, 通过设置垂直样带, 同时结合区域温度、降水的长时间序列分析, 探究两处树线的时空动态过程, 并明确了建群种冷杉(Abies spp.)的生态学特征。结果表明: 1)折多山和剪子弯山区域的气温在过去58年均存在显著的上升趋势(分别上升了0.72和0.91 ℃), 而折多山和剪子弯山区域降水均存在微弱的降低趋势。2)折多山的峨眉冷杉(A. fabri)龄级结构呈反J形, 剪子弯山的鳞皮冷杉(A. squamata)龄级结构呈双峰形, 二者种群结构均相对稳定。3)在小尺度上, 种子扩散限制使得两处树线的冷杉聚集分布。在大尺度上, 折多山峨眉冷杉亦呈聚集分布, 而剪子弯山鳞皮冷杉受生长环境以及种内或种间关系的影响呈随机分布。4)两处样地建群树种的树高和基径均随海拔升高而降低, 位于树线交错带上部的冷杉均呈现树高生长大于径向生长的异速生长关系, 而位于样地中、下部位的冷杉大部分呈等速生长关系。5)相比10年前, 折多山和剪子弯山的树线及树种线位置均无明显变化, 剪子弯山鳞皮冷杉种群的树木密度亦无明显变化, 而折多山的树木个体数提高了约25%; 相比20年前, 折多山和剪子弯山的树种线分别上移了50和30 m, 树线位置分别升高了75和40 m, 树木个体数亦明显增加, 分别提高了约220%和100%。树线及其建群种在较大时空尺度上主要受热量的控制, 而在较小时空尺度上受温度及生长环境共同作用的影响。
周天阳, NARAYAN Prasad Gaire, 廖礼彬, 郑莉莉, 王金牛, 孙建, 魏彦强, 谢雨, 吴彦. 青藏高原东缘两处高山树线交错带时空动态及其建群种的生态学特征. 植物生态学报, 2018, 42(11): 1082-1093. DOI: 10.17521/cjpe.2018.0082
ZHOU Tian-Yang, NARAYAN Prasad Gaire, LIAO Li-Bin, ZHENG Li-Li, WANG Jin-Niu, SUN Jian, WEI Yan-Qiang, XIE Yu, WU Yan. Spatio-temporal dynamics of two alpine treeline ecotones and ecological characteristics of their dominate species at the eastern margin of Qinghai-Xizang Plateau. Chinese Journal of Plant Ecology, 2018, 42(11): 1082-1093. DOI: 10.17521/cjpe.2018.0082
| 样地 Plot | 海拔 Altitude (m) | 经纬度 Longitude and latitude | 坡向 Slope aspect | 坡度 Slope | 主要树种 Main species |
|---|---|---|---|---|---|
| 折多山 Zheduo Mountain | 4 210-4 380 | 101.73° E, 30.05° N | 正北 North | 27.5° | 峨眉冷杉、川西云杉、四川红杉 Abies fabri, Picea likiangensis, Larix mastersiana |
| 剪子弯山 Jianziwan Mountain | 4 360-4 520 | 100.82° E, 30.02° N | 西偏北30° North-west 30° | 25.3° | 鳞皮冷杉、四川红杉 Abies squamata, Larix mastersiana |
表1 折多山和剪子弯山树线交错带样地概况
Table 1 The basic information of treeline ecotone of Zheduo Mountain and Jianziwan Mountain
| 样地 Plot | 海拔 Altitude (m) | 经纬度 Longitude and latitude | 坡向 Slope aspect | 坡度 Slope | 主要树种 Main species |
|---|---|---|---|---|---|
| 折多山 Zheduo Mountain | 4 210-4 380 | 101.73° E, 30.05° N | 正北 North | 27.5° | 峨眉冷杉、川西云杉、四川红杉 Abies fabri, Picea likiangensis, Larix mastersiana |
| 剪子弯山 Jianziwan Mountain | 4 360-4 520 | 100.82° E, 30.02° N | 西偏北30° North-west 30° | 25.3° | 鳞皮冷杉、四川红杉 Abies squamata, Larix mastersiana |
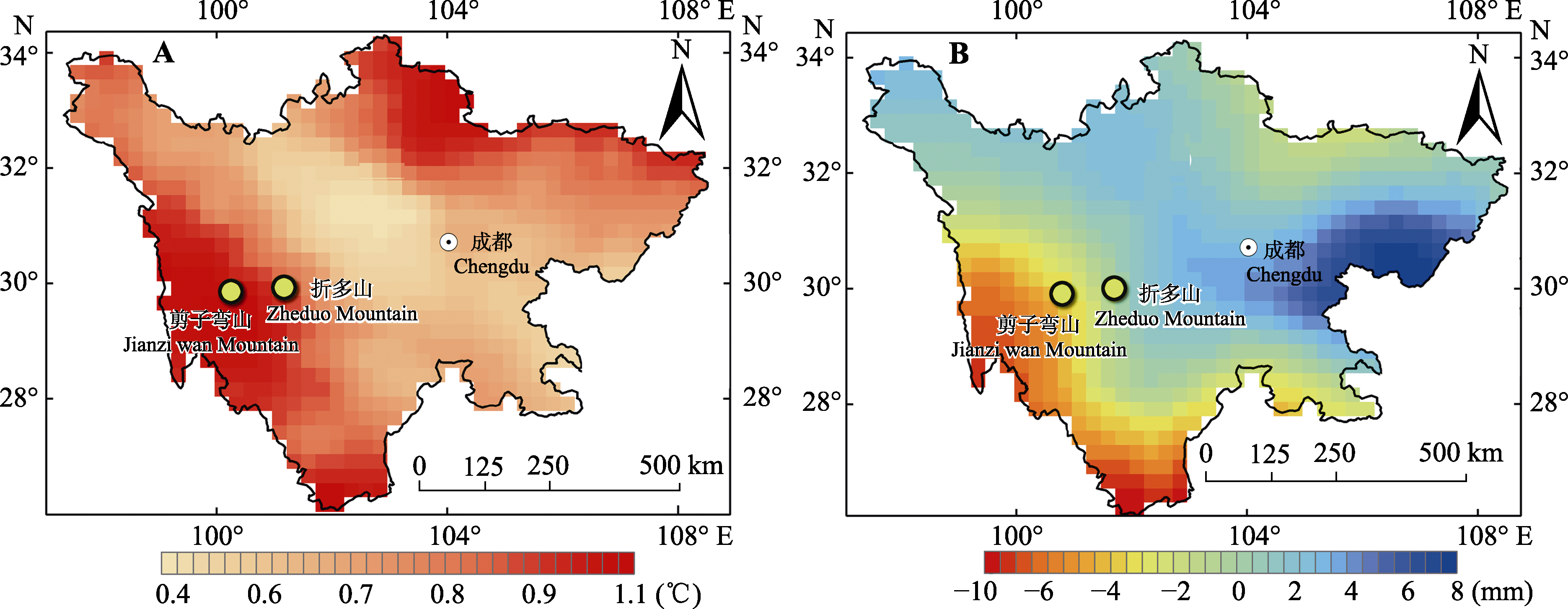
图1 2010-2017年与1961-1970年的差值。A, 年平均气温。B, 年降水量。
Fig. 1 The difference value map between 2010-2017 and 1961-1970. A, Annual mean temperature. B, Annual precipitation.
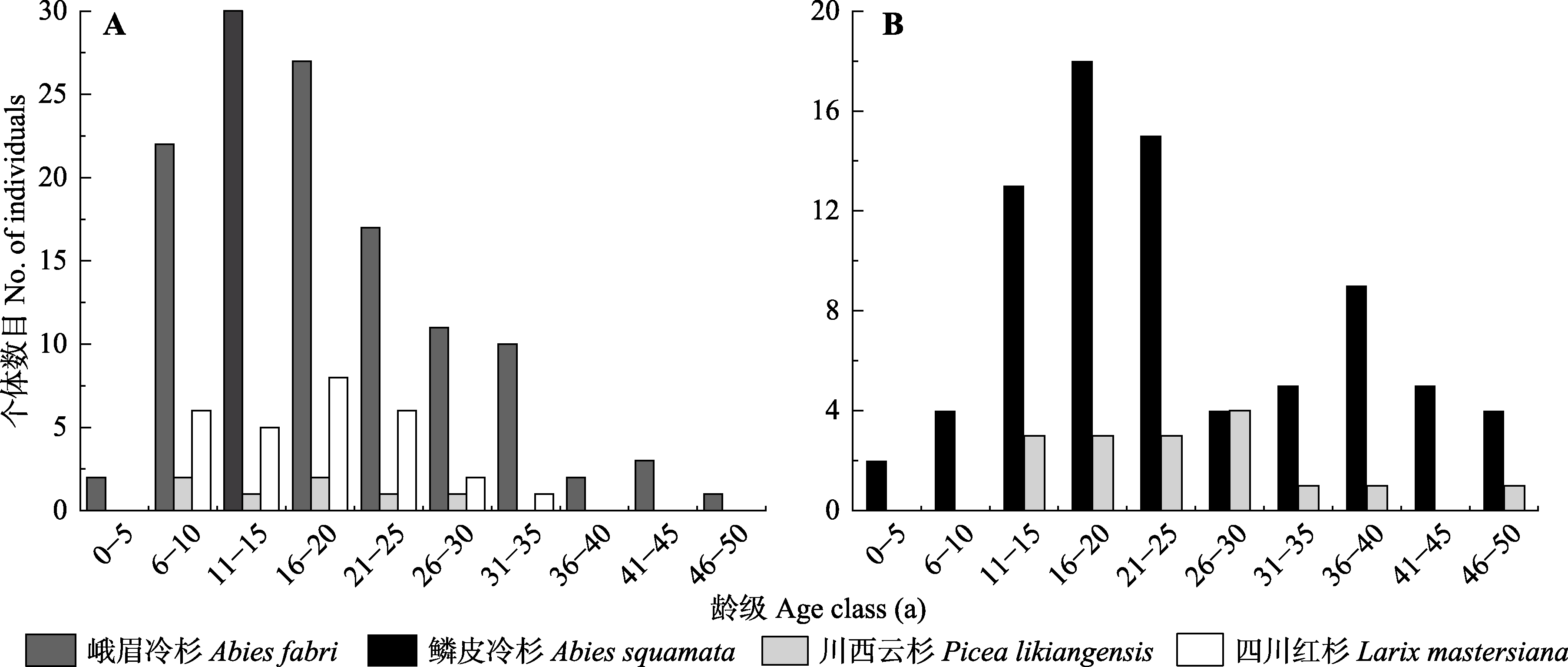
图2 折多山(A)及剪子弯山(B)样地树种龄级结构。
Fig. 2 Age structure of treeline-forming species at treeline in the Zheduo Mountain (A) and Jianziwan Mountain (B).
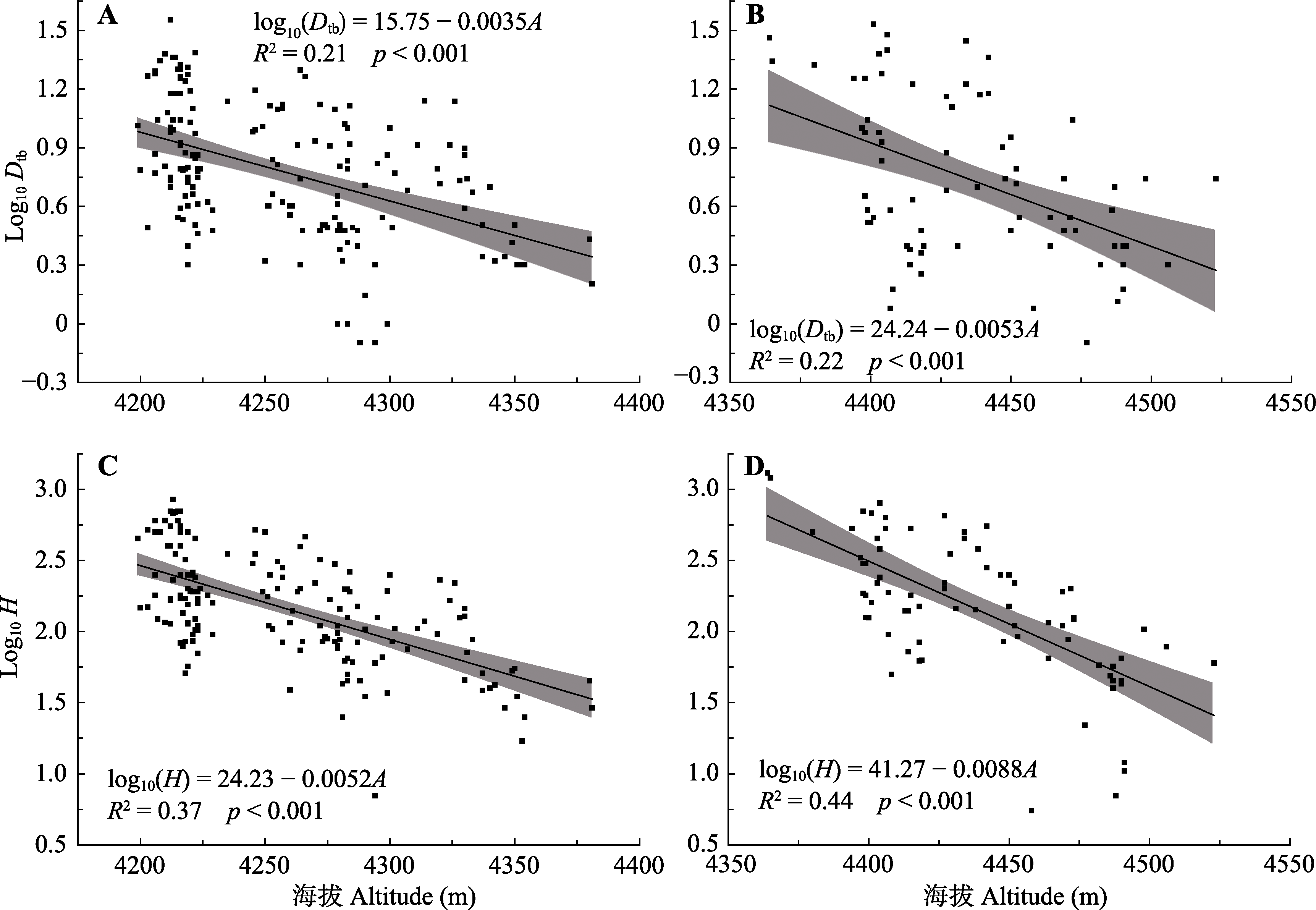
图3 折多山(A, C)与剪子弯山(B, D)的树高(H)、基径(Dtb)与海拔(A)的关系。
Fig. 3 The relationships of tree height(H), base diameter (Dtb) and altitude (A) in Zheduo Mountain (A, C) and Jianziwan Mountain(B, D).
| 样点 Plot | Y轴 Y (m) | R2 | p | 斜率(95%置信区间) Slope (95% confidence interval) | 等速生长检验 Test of isometry p |
|---|---|---|---|---|---|
| 折多山 Zheduo Mountain | <100 | 0.80 | <0.001 | 1.64 (1.30, 2.08)a | <0.001 |
| 100-200 | 0.54 | <0.001 | 0.72 (0.51, 1.00)c | 0.053 | |
| 201-300 | 0.72 | <0.001 | 1.15 (0.98, 1.34)b | 0.08 | |
| >300 | 0.52 | <0.001 | 1.12 (0.90, 1.34)b | 0.296 | |
| 剪子弯山 Jianziwan Mountain | <100 | 0.75 | <0.001 | 1.53 (1.21, 1.93)a | <0.001 |
| 100-200 | 0.85 | <0.001 | 0.99 (0.80, 1.22)b | 0.92 | |
| 201-300 | 0.80 | <0.001 | 0.83 (0.63, 1.10)b | 0.18 | |
| >300 | 0.90 | <0.001 | 0.85 (0.74, 0.98)b | 0.028 |
表2 折多山与剪子弯山冷杉径向生长与高生长间的关系
Table 2 The correlation between the growth of height and base diameter in Zhedou Mountain and Jianziwan Mountain
| 样点 Plot | Y轴 Y (m) | R2 | p | 斜率(95%置信区间) Slope (95% confidence interval) | 等速生长检验 Test of isometry p |
|---|---|---|---|---|---|
| 折多山 Zheduo Mountain | <100 | 0.80 | <0.001 | 1.64 (1.30, 2.08)a | <0.001 |
| 100-200 | 0.54 | <0.001 | 0.72 (0.51, 1.00)c | 0.053 | |
| 201-300 | 0.72 | <0.001 | 1.15 (0.98, 1.34)b | 0.08 | |
| >300 | 0.52 | <0.001 | 1.12 (0.90, 1.34)b | 0.296 | |
| 剪子弯山 Jianziwan Mountain | <100 | 0.75 | <0.001 | 1.53 (1.21, 1.93)a | <0.001 |
| 100-200 | 0.85 | <0.001 | 0.99 (0.80, 1.22)b | 0.92 | |
| 201-300 | 0.80 | <0.001 | 0.83 (0.63, 1.10)b | 0.18 | |
| >300 | 0.90 | <0.001 | 0.85 (0.74, 0.98)b | 0.028 |
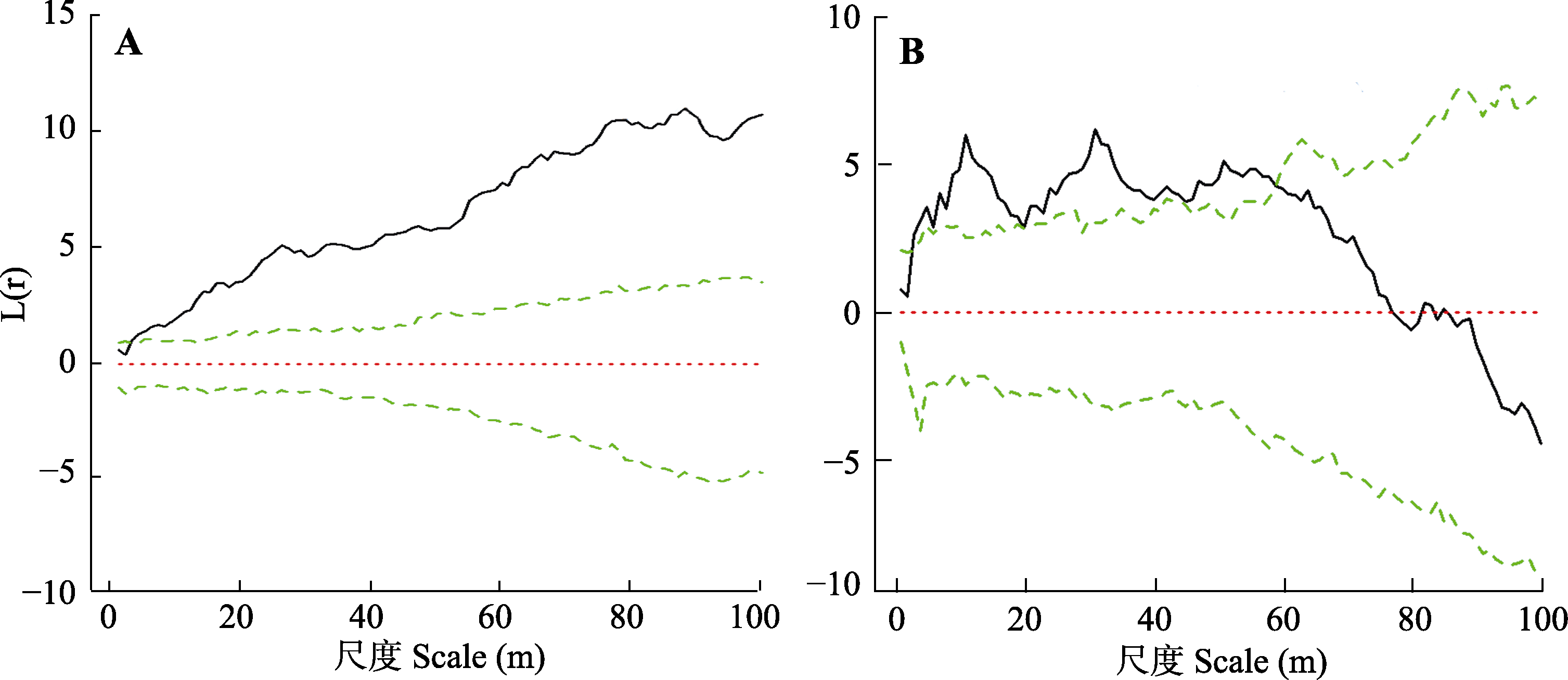
图4 折多山(A)及剪子弯山(B)样地点空间格局分析。两条绿线为拟合的置信区间, 黑线为模拟结果。
Fig. 4 Point pattern analyses for the Zheduo Mountain (A) and Jianziwan Mountain (B). The two green lines are the confidence intervals for the fitting, and the solid line is from the simulation.
| 样地 Plot | 回归方程 Regression equation | R2 | p |
|---|---|---|---|
| 折多山 Zheduo Mountain | H = 34.80e0.079a | 0.68 | < 0.001 |
| 剪子弯山 Jianziwan Mountain | H = 14.38e0.087a | 0.71 | < 0.001 |
表3 折多山与剪子弯山树高(H)与树龄(a)的回归模型
Table 3 The regression model of tree height (H) and tree age (a)
| 样地 Plot | 回归方程 Regression equation | R2 | p |
|---|---|---|---|
| 折多山 Zheduo Mountain | H = 34.80e0.079a | 0.68 | < 0.001 |
| 剪子弯山 Jianziwan Mountain | H = 14.38e0.087a | 0.71 | < 0.001 |
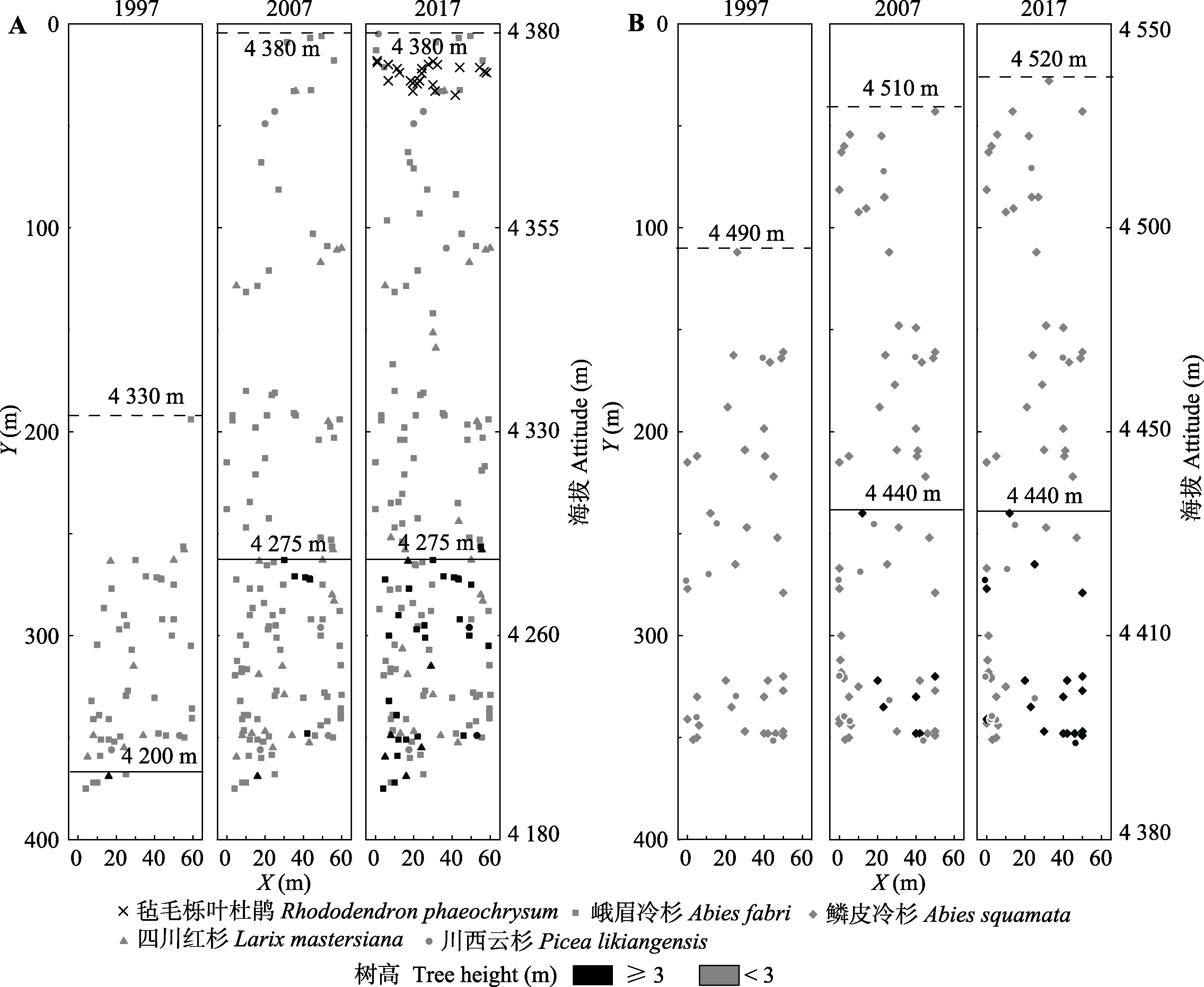
图5 折多山(A)及剪子弯山(B)样地树线时空模拟。图中黑线为树线, 虚线为树种线。
Fig. 5 The spatio-temporal pattern of treeline dynamics in the Zheduo Mountain (A) and Jianziwan Mountain (B). The solid line is the position of treeline and the dash line is tree species line.
| [30] |
Luckman B, Kavanagh T ( 2000). Impact of climate fluctuations on mountain environments in the Canadian Rockies. AMBIO, 29, 371-380.
DOI URL |
| [31] |
MacDonald GM, Szeicz JM, Claricoates J, Dale KA ( 1998). Response of the central Canadian treeline to recent climatic changes. Annals of the Association of American Geographers, 88, 183-208.
DOI URL |
| [32] |
Mayr S, Charra-Vaskou K ( 2007). Winter at the alpine timberline causes complex within-tree patterns of water potential and embolism in Picea abies. Physiologia Plantarum, 131, 131-139.
DOI URL |
| [33] |
Niklas KJ ( 2005). Modelling below- and above-ground biomass for non-woody and woody plants. Annals of Botany, 95, 315-321.
DOI URL PMID |
| [34] | Pelissier R, Goreaud F ( 2015). Ads package for R: A fast unbiased implementation of the k-function family for studying spatial point patterns in irregular-shaped sampling windows. Journal of Statistical Software, 63, 1-18. |
| [35] |
Ran F, Liang YM, Yang Y, Yang Y, Wang GX ( 2014). Spatial-temporal dynamics of an Abies fabri population near the alpine treeline in the Yajiageng area of Gongga Mountain, China. Acta Ecologica Sinica, 34, 6872-6878.
DOI URL |
|
[ 冉飞, 梁一鸣, 杨燕, 杨阳, 王根绪 ( 2014). 贡嘎山雅家埂峨眉冷杉林线种群的时空动态. 生态学报, 34, 6872-6878.]
DOI URL |
|
| [36] |
Ripley BD ( 1977). Modeling spatial patterns. Journal of the Royal Statistical Society Series B—Methodological, 39, 172-212.
DOI URL |
| [37] |
Shi CM, Masson-Delmotte V, Daux V, Li ZS, Carre M, Moore JC ( 2015). Unprecedented recent warming rate and temperature variability over the east Tibetan Plateau inferred from alpine treeline dendrochronology. Climate Dynamics, 45, 1367-1380.
DOI URL |
| [38] |
Shi JY, Han HR, Cheng XQ, Dong LL, Tian HX, Cai MK, Kang FF ( 2017). Age structure and dynamics of Pinus tabuliformis population in the Liaoheyuan Nature Reserve of Hebei Province. Chinese Journal of Ecology, 36, 1808-1814.
DOI URL |
|
[ 矢佳昱, 韩海荣, 程小琴, 董玲玲, 田慧霞, 蔡锰柯, 康峰峰 ( 2017). 河北辽河源自然保护区油松种群年龄结构和种群动态. 生态学杂志, 36, 1808-1814.]
DOI URL |
|
| [39] |
Stevens GC, Fox JF ( 1991). The causes of treeline. Annual Review of Ecology and Systematics, 22, 177-191.
DOI URL |
| [40] |
Stewart GH, Rose AB ( 1990). The significance of life-history strategies in the developmental history of mixed beech (nothofagus) forests, New-Zealand. Vegetatio, 87, 101-114.
DOI URL |
| [41] |
van Bogaert R, Haneca K, Hoogesteger J, Jonasson C, de Dapper M, Callaghan TV ( 2011). A century of tree line changes in sub-Arctic Sweden shows local and regional variability and only a minor influence of 20th Century climate warming. Journal of Biogeography, 38, 907-921.
DOI URL |
| [42] |
Wang T, Zhang QB, Ma KP ( 2006). Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China. Global Ecology and Biogeography, 15, 406-415.
DOI URL |
| [43] |
Wang YF, Julio Camarero J, Luo TX, Liang EY ( 2012). Spatial patterns of smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns. Plant Ecology & Diversity, 5, 311-321.
DOI URL |
| [44] | Wang YF, Liang EY ( 2012). A review on progresses in treeline dynamics and climate change. Journal of Earth Environment, 3, 855-861. |
| [ 王亚锋, 梁尔源 ( 2012). 树线波动与气候变化研究进展. 地球环境学报, 3, 855-861.] | |
| [45] |
Wang YF, Liang EY, Lu XM, Zhu HF, Piao SL, Zhu LP ( 2017). Are treelines advancing in response to climate warming on the Tibetan Plateau? Chinese Journal of Nature, 39, 179-183.
DOI URL |
|
[ 王亚锋, 梁尔源, 芦晓明, 朱海峰, 朴世龙, 朱立平 ( 2017). 气候变暖会使青藏高原树线一直上升吗? 自然杂志, 39, 179-183.]
DOI URL |
|
| [46] |
Warton DI, Duursma RA, Falster DS, Taskinen S ( 2012). Smatr 3—An R package for estimation and inference about allometric lines. Methods in Ecology and Evolution, 3, 257-259.
DOI URL |
| [47] |
Warton DI, Wright IJ, Falster DS, Westoby M ( 2006). Bivariate line-fitting methods for allometry. Biological Reviews, 81, 259-291.
DOI URL PMID |
| [48] |
Wilmking M, Juday GP, Barber VA, Zald HSJ ( 2004). Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds. Global Change Biology, 10, 1724-1736.
DOI URL |
| [49] |
Wong MH, Duan CQ, Long YC, Luo YM, Xie GQ ( 2010). How will the distribution and size of subalpine Abies georgei forest respond to climate change? A study in northwest Yunnan, China. Physical Geography, 31, 319-335.
DOI URL |
| [50] | Xie CQ, Tian MX, Zhao ZR, Zheng WL, Wang GY ( 2015). Spatial point pattern analysis of Abies georgei var. smithii in forest of Sygera Mountains in southeast Tibet, China. Chinese Journal of Applied Ecology, 26, 1617-1624. |
| [ 解传奇, 田民霞, 赵忠瑞, 郑维列, 王国严 ( 2015). 西藏色季拉山急尖长苞冷杉种群点格局分析. 应用生态学报, 26, 1617-1624.] | |
| [51] |
Xie ZQ, Chen WL, Lu P, Hu D ( 1999). The demography and age structure of the endangered plant population of Cathaya argyrophylla. Acta Ecologica Sinica, 19, 523-528.
DOI URL |
|
[ 谢宗强, 陈伟烈, 路鹏, 胡东 ( 1999). 濒危植物银杉的种群统计与年龄结构. 生态学报, 19, 523-528.]
DOI URL |
|
| [52] |
Yang H, Li YL, Shen L, Kang XG, Yue G, Wang Y ( 2014). Spatial distribution patterns of seedling and sapling in a spruce-fir forest in the Changbai Mountains area in northeastern China. Acta Ecologica Sinica, 34, 7311-7319.
DOI URL |
|
[ 杨华, 李艳丽, 沈林, 亢新刚, 岳刚, 王妍 ( 2014). 长白山云冷杉林幼苗幼树空间分布格局及其更新特征. 生态学报, 34, 7311-7319.]
DOI URL |
|
| [53] |
Yang XD, Yan ER, Zhang ZH, Sun BW, Huang HX, Arshad A, Ma WJ, Shi QR ( 2013). Tree architecture of overlapping species among successional stages in evergreen broad-leaved forests in Tiantong region, Zhejiang Province, China. Chinese Journal of Plant Ecology, 37, 611-619.
DOI URL |
|
[ 杨晓东, 阎恩荣, 张志浩, 孙宝伟, 黄海侠, Arshad A, 马文济, 史青茹 ( 2013). 浙江天童常绿阔叶林演替阶段共有种的树木构型. 植物生态学报, 37, 611-619.]
DOI URL |
|
| [1] | Bai DZ ( 2012). The Impact Factors of Growth and Regeneration of Picea crassifolia Growing at Timberline in The Qilian Mountains. PhD dissertation, Chinese Academy of Forestry, Beijing. |
| [ 白登忠 ( 2012). 祁连山青海云杉林线树木生长、更新的影响因素研究. 博士学位论文, 中国林业科学研究院, 北京.] | |
| [54] | Zhang JT ( 1998). Analysis of spatial point pattern for plant species. Acta Phytocologica Sinica, 22, 57-62. |
| [ 张金屯 ( 1998). 植物种群空间分布的点格局分析. 植物生态学报, 22, 57-62.] | |
| [2] |
Batllori E, Blanco-Moreno JM, Ninot JM, Gutierrez E, Carrillo E ( 2009). Vegetation patterns at the alpine treeline ecotone: The influence of tree cover on abrupt change in species composition of alpine communities. Journal of Vegetation Science, 20, 814-825.
DOI URL |
| [3] |
Camarero JJ, Gutierrez E ( 2004). Pace and pattern of recent treeline dynamics, response of ecotones to climatic variability in the Spanish Pyrenees. Climatic Change, 63, 181-200.
DOI URL |
| [55] |
Zhang PJ, Qing H, Zhang L, Xu YD, Mu L, Ye RH, Qiu X, Chang H, Shen HH, Yang J ( 2017). Population structure and spatial pattern of Caragana tibetica communities in Nei Mongol shrub-encroached grassland. Chinese Journal of Plant Ecology, 41, 165-174.
DOI URL |
|
[ 张璞进, 清华, 张雷, 徐延达, 木兰, 晔薷罕, 邱晓, 常虹, 沈海花, 杨劼 ( 2017). 内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局. 植物生态学报, 41, 165-174.]
DOI URL |
|
| [4] |
Cheng W, Luo P, Wu N ( 2005). Ecological characteristics of minjiang fir (Abies faxoniana Rehd. et Wild) population near timberline on upper Min River. Chinese Journal of Applied and Envirnmental Biology, 11, 300-303.
DOI URL |
|
[ 程伟, 罗鹏, 吴宁 ( 2005). 岷江上游林线附近岷江冷杉种群(Abies faxoniana Rehd. et Wild)的生态学特点. 应用与环境生物学报, 11, 300-303.]
DOI URL |
|
| [5] |
Condit R, Ashton PS, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell SP, Foster RB, Itoh A, Lafrankie JV, Lee HS, Losos E, Manokaran N, Sukumar R, Yamakura T ( 2000). Spatial patterns in the distribution of tropical tree species. Science, 288, 1414-1418.
DOI URL PMID |
| [6] |
Dai JH, Cui HT ( 1999). A reviewon the studies of alpine timberline. Scientia Geographica Sinica, 19(3), 52-58.
DOI URL |
|
[ 戴君虎, 崔海亭 ( 1999). 国内外高山林线研究综述. 地理科学, 19(3), 52-58.]
DOI URL |
|
| [7] |
Falster DS, Westoby M ( 2003). Plant height and evolutionary games. Trends in Ecology & Evolution, 18, 337-343.
DOI URL |
| [8] |
Gaire NP, Koirala M, Bhuju DR, Borgaonkar HP ( 2014). Treeline dynamics with climate change at the Central Nepal Himalaya. Climate of the Past, 10, 1277-1290.
DOI URL |
| [9] |
Gao J, Wang JN, Xu B, Xie Y, He JD, Wu Y ( 2016). Plant leaf traits, height and biomass partitioning in typical ephemerals under different levels of snow cover thickness in an alpine meadow. Chinese Journal of Plant Ecology, 40, 775-787.
DOI URL |
|
[ 高景, 王金牛, 徐波, 谢雨, 贺俊东, 吴彦 ( 2016). 不同雪被厚度下典型高山草地早春植物叶片性状、株高及生物量分配的研究. 植物生态学报, 40, 775-787.]
DOI URL |
|
| [10] |
Harsch MA, Hulme PE, Mcglone MS, Duncan RP ( 2009). Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecology Letters, 12, 1040-1049.
DOI URL |
| [11] | IPCC (Intergovernmental Panel on Climate Change) (2014). Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. |
| [12] |
Jin HJ, Ma QL, Zhang DK, Liu YJ, Yuan HB ( 2012). Analysis on typical shrub plant community characteristics and quantitative characteristics in Ulanbuh desert. Acta Botanica Boreall-Occidentalia Sinica, 32, 579-588.
DOI URL |
|
[ 靳虎甲, 马全林, 张德魁, 刘有军, 袁洪波 ( 2012). 乌兰布和沙漠典型灌木群落结构及数量特征. 西北植物学报, 32, 579-588.]
DOI URL |
|
| [13] |
Kim JW, Lee JS ( 2015). Dynamics of alpine treelines: Positive feedbacks and global, regional and local controls. Journal of Ecology and Environment, 38, 1-14.
DOI URL |
| [14] |
Klasner FL, Fagre DB ( 2002). A half century of change in alpine treeline patterns at Glacier National Park, Montana, USA. Arctic Antarctic and Alpine Research, 34, 49-56.
DOI URL |
| [15] |
Knowles P, Grant MC ( 1983). Age and size structure analyses of engelmann spruce, ponderosa pine, lodgepole pine, and limber pine in Colorado. Ecology, 64, 1-9.
DOI URL |
| [16] |
Körner C ( 1998). A re-assessment of high elevation treeline positions and their explanation. Oecologia, 115, 445-459.
DOI URL PMID |
| [17] | Körner C ( 2003). Alpine Plant Life. 2nd edn. Springer, Berlin. |
| [18] | Körner C (Translated by Wu N, Shi PL, Yi SL, Wang JN)( 2017). Alpine Treelines. Publishing House of Electronics Industry, Beijing. |
| [吴宁,石培礼,易绍良, 王金牛 (译)( 2017). 高山树线——全球高海拔树木生长上线的功能生态学. 电子工业出版社, 北京.] | |
| [19] |
Körner C, Paulsen J ( 2004). A world-wide study of high altitude treeline temperatures. Journal of Biogeography, 31, 713-732.
DOI URL |
| [20] | Kullman L ( 1993). Tree timit dynamics of Betula pubescens ssp. tortuosa in relation to climate variability: Evidence from Central Sweden. Journal of Vegetation Science, 4, 765. |
| [21] |
Kullman L ( 2001). 20th century climate warming and tree-limit rise in the southern Scandes of Sweden. AMBIO, 30, 72-80.
DOI URL PMID |
| [22] |
Lan GY, Hu YH, Cao M, Zhu H, Wang H, Zhou SS, Deng XB, Cui JY, Huang JG, Liu LY, Xu HL, Song JP, He YC ( 2008). Establishment of Xishuangbanna tropical forest dynamics plot: Species compositions and spatial distribution patterns. Chinese Journal of Plant Ecology (Chinese Version), 32, 287-298.
DOI URL |
|
[ 兰国玉, 胡跃华, 曹敏, 朱华, 王洪, 周仕顺, 邓晓保, 崔景云, 黄建国, 刘林云, 许海龙, 宋军平, 何有才 ( 2008). 西双版纳热带森林动态监测样地——树种组成与空间分布格局. 植物生态学报, 32, 287-298.]
DOI URL |
|
| [23] |
Li GC, Song HD, Li Q, Bu SH ( 2017). Spatial point pattern analysis of main trees and flowering Fargesia qinlingensis in Abies fargesii forests in Mt. Taibai of the Qinling Mountains, China. Chinese Journal of Applied Ecology, 28, 3487-3493.
DOI URL |
|
[ 李国春, 宋华东, 李琦, 卜书海 ( 2017). 太白山巴山冷杉林主要树种与开花秦岭箭竹的空间点格局分析. 应用生态学报, 28, 3487-3493.]
DOI URL |
|
| [24] | Li LP, Mohamma A, Wang XP ( 2011). Study on relationship between height and DBH of mountain coniferous forests in Xinjiang. Arid Zone Research, 28(1), 47-53. |
| [ 李利平, 安尼瓦尔·买买提, 王襄平 ( 2011). 新疆山地针叶林乔木胸径-树高关系分析. 干旱区研究, 28(1), 47-53.] | |
| [25] |
Liang EY, Leuschner C, Dulamsuren C, Wagner B, Hauck M ( 2016). Global warming-related tree growth decline and mortality on the north-eastern Tibetan Plateau. Climatic Change, 13, 163-176.
DOI URL |
| [26] |
Liang EY, Wang YF, Eckstein D, Luo TX ( 2011). Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytologist, 190, 760-769.
DOI URL |
| [27] |
Liu XD, Chen BD ( 2000). Climatic warming in the Tibetan Plateau during recent decades. International Journal of Climatology, 20, 1729-1742.
DOI URL |
| [28] |
Lloyd AH, Fastie CL ( 2002). Spatial and temporal variability in the growth and climate response of treeline trees in Alaska. Climate Change, 52, 481-509.
DOI URL |
| [29] |
Lloyd AH, Fastie CL ( 2003). Recent changes in treeline forest distribution and structure in interior Alaska. Ecoscience, 10, 176-185.
DOI URL |
| [1] | 张尧, 陈岚, 王洁莹, 李益, 王俊, 郭垚鑫, 任成杰, 白红英, 孙昊田, 赵发珠. 太白山不同海拔森林根际土壤微生物碳利用效率差异性及其影响因素[J]. 植物生态学报, 2023, 47(2): 275-288. |
| [2] | 刘艳杰, 刘玉龙, 王传宽, 王兴昌. 东北温带森林5个羽状复叶树种叶成本-效益关系比较[J]. 植物生态学报, 2023, 47(11): 1540-1550. |
| [3] | 王广亚, 陈柄华, 黄雨晨, 金光泽, 刘志理. 着生位置对水曲柳小叶性状变异及性状间相关性的影响[J]. 植物生态学报, 2022, 46(6): 712-721. |
| [4] | 李露, 金光泽, 刘志理. 阔叶红松林3种阔叶树种柄叶性状变异与相关性[J]. 植物生态学报, 2022, 46(6): 687-699. |
| [5] | 卢晶, 马宗祺, 高鹏斐, 樊宝丽, 孙坤. 祁连山区演替先锋物种西藏沙棘的种群结构及动态对海拔梯度的响应[J]. 植物生态学报, 2022, 46(5): 569-579. |
| [6] | 熊映杰, 于果, 魏凯璐, 彭娟, 耿鸿儒, 杨冬梅, 彭国全. 天童山阔叶木本植物叶片大小与叶脉密度及单位叶脉长度细胞壁干质量的关系[J]. 植物生态学报, 2022, 46(2): 136-147. |
| [7] | 李东, 田秋香, 赵小祥, 林巧玲, 岳朋芸, 姜庆虎, 刘峰. 贡嘎山树线过渡带土壤胞外酶活性及其化学计量比特征[J]. 植物生态学报, 2022, 46(2): 232-242. |
| [8] | 牟文博, 徐当会, 王谢军, 敬文茂, 张瑞英, 顾玉玲, 姚广前, 祁世华, 张龙, 苟亚飞. 排露沟流域不同海拔灌丛土壤碳氮磷化学计量特征[J]. 植物生态学报, 2022, 46(11): 1422-1431. |
| [9] | 董楠, 唐明明, 崔文倩, 岳梦瑶, 刘洁, 黄玉杰. 不同根系分隔方式对栗和茶幼苗生长的影响[J]. 植物生态学报, 2022, 46(1): 62-73. |
| [10] | 尹晓雷, 刘旭阳, 金强, 李先德, 林少颖, 阳祥, 王维奇, 张永勋. 不同管理模式对茶树碳氮磷含量及其生态化学计量比的影响[J]. 植物生态学报, 2021, 45(7): 749-759. |
| [11] | 杨克彤, 常海龙, 陈国鹏, 俞筱押, 鲜骏仁. 兰州市主要绿化植物气孔性状特征[J]. 植物生态学报, 2021, 45(2): 187-196. |
| [12] | 解梦怡, 冯秀秀, 马寰菲, 胡汗, 王洁莹, 郭垚鑫, 任成杰, 王俊, 赵发珠. 秦岭锐齿栎林土壤酶活性与化学计量比变化特征及其影响因素[J]. 植物生态学报, 2020, 44(8): 885-894. |
| [13] | 邢磊, 段娜, 李清河, 刘成功, 李慧卿, 孙高洁. 白刺不同物候期的生物量分配规律[J]. 植物生态学报, 2020, 44(7): 763-771. |
| [14] | 熊星烁, 蔡宏宇, 李耀琪, 马文红, 牛克昌, 陈迪马, 刘娜娜, 苏香燕, 景鹤影, 冯晓娟, 曾辉, 王志恒. 内蒙古典型草原植物叶片碳氮磷化学计量特征的季节动态[J]. 植物生态学报, 2020, 44(11): 1138-1153. |
| [15] | 陈国鹏, 杨克彤, 王立, 王飞, 曹秀文, 陈林生. 甘肃南部7种高寒杜鹃生物量分配的异速生长关系[J]. 植物生态学报, 2020, 44(10): 1040-1049. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19