
植物生态学报 ›› 2017, Vol. 41 ›› Issue (1): 95-104.DOI: 10.17521/cjpe.2015.0302
所属专题: 中国灌丛生态系统碳储量的研究; 全球变化与生态系统; 生态系统碳水能量通量; 土壤呼吸
张蔷1,2, 李家湘3, 谢宗强1,*( )
)
收稿日期:2015-08-17
接受日期:2016-01-17
出版日期:2017-01-10
发布日期:2017-01-23
通讯作者:
谢宗强
作者简介:* 通信作者Author for correspondence (E-mail:基金资助:
Qiang ZHANG1,2, Jia-Xiang LI3, Zong-Qiang XIE1,*()
Received:2015-08-17
Accepted:2016-01-17
Online:2017-01-10
Published:2017-01-23
Contact:
Zong-Qiang XIE
About author:KANG Jing-yao(1991-), E-mail: 摘要:
为探究灌丛生态系统对大气氮沉降的响应, 2013年1月至2014年9月, 对湖南大围山杜鹃(Rhododendron simsii)灌丛群落进行了短期模拟氮沉降试验, 施氮浓度分别为0 (CK)、2 (LN)、5 (MN)和10 (HN) g·m-2·a-1。利用LI-8100土壤碳通量测量系统测定土壤呼吸速率, 并测定不同氮处理下根系生物量增量和凋落物量。结果表明: 该地区土壤呼吸呈现明显的季节动态, 夏季土壤呼吸最强, 冬季最弱。CK、LN、MN和HN处理样地每年通过土壤呼吸释放的CO2量分别为2.37、2.79、2.26和2.30 kg CO2·m-2。CK、LN、MN和HN处理下, 年平均土壤呼吸速率分别为1.71、2.01、1.63和1.66 μmol CO2·m-2·s-1, LN处理样地的年均土壤呼吸速率与对照样地相比增加了17.25%, MN和HN处理则比对照样地稍低。施氮增加了根系生物量增量和凋落物量, 但没有达到显著水平。土壤呼吸速率与5 cm土壤温度呈显著指数相关关系, 与5 cm土壤的含水量呈显著线性相关关系。CK、LN、MN和HN处理下, 土壤呼吸的温度敏感性(Q10)值分别为3.96、3.60、3.71和3.51, 表明施氮降低了温度敏感性。氮添加导致的根系生物量增加是引起该区域土壤呼吸速率变化的一个重要原因。
张蔷, 李家湘, 谢宗强. 氮添加对亚热带山地杜鹃灌丛土壤呼吸的影响. 植物生态学报, 2017, 41(1): 95-104. DOI: 10.17521/cjpe.2015.0302
Qiang ZHANG, Jia-Xiang LI, Zong-Qiang XIE. Effects of nitrogen addition on soil respiration of Rhododendron simsii shrubland in the subtropical mountainous areas of China. Chinese Journal of Plant Ecology, 2017, 41(1): 95-104. DOI: 10.17521/cjpe.2015.0302
| 土壤深度 Soil depth (cm) | 全碳 Total C (%) | 全氮 Total N (%) | C:N | 全磷 Total P (mg·g-1) | pH | 土壤容重 Soil bulk density (g·cm-3) |
|---|---|---|---|---|---|---|
| 0-10 | 4.22 ± 0.16a | 0.36 ± 0.01a | 11.63 ± 0.14a | 0.77 ± 0.02a | 5.11 ± 0.01b | 0.88 ± 0.01b |
| 10-20 | 2.72 ± 0.13b | 0.26 ± 0.01b | 10.42 ± 0.19b | 0.75 ± 0.02a | 5.17 ± 0.02b | 0.98 ± 0.02a |
| 20-30 | 2.03 ± 0.13c | 0.21 ± 0.01c | 9.48 ± 0.25c | 0.75 ± 0.03a | 5.31 ± 0.03a | 1.01 ± 0.03a |
表1 不同土层的土壤理化性质(平均值±标准误差)
Table 1 Soil physical and chemical properties at different depth (mean ± SE)
| 土壤深度 Soil depth (cm) | 全碳 Total C (%) | 全氮 Total N (%) | C:N | 全磷 Total P (mg·g-1) | pH | 土壤容重 Soil bulk density (g·cm-3) |
|---|---|---|---|---|---|---|
| 0-10 | 4.22 ± 0.16a | 0.36 ± 0.01a | 11.63 ± 0.14a | 0.77 ± 0.02a | 5.11 ± 0.01b | 0.88 ± 0.01b |
| 10-20 | 2.72 ± 0.13b | 0.26 ± 0.01b | 10.42 ± 0.19b | 0.75 ± 0.02a | 5.17 ± 0.02b | 0.98 ± 0.02a |
| 20-30 | 2.03 ± 0.13c | 0.21 ± 0.01c | 9.48 ± 0.25c | 0.75 ± 0.03a | 5.31 ± 0.03a | 1.01 ± 0.03a |
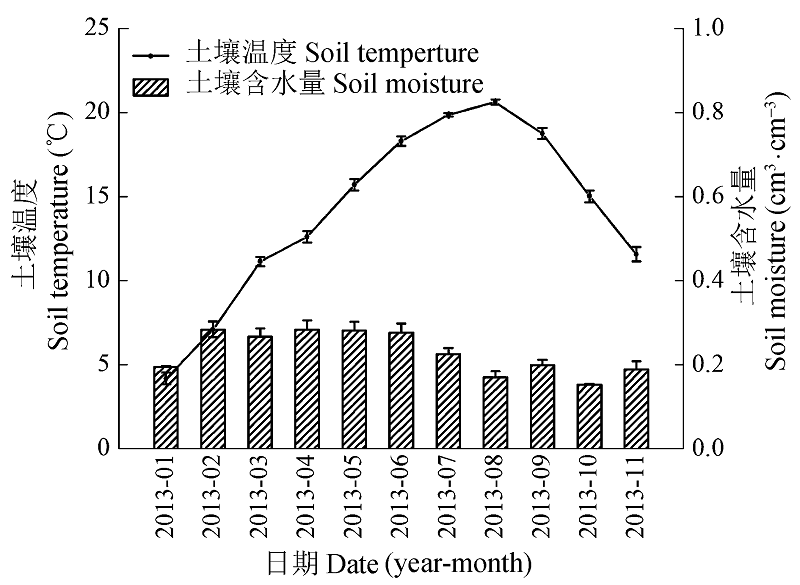
图1 土壤温度和含水量的季节变化(平均值±标准误差)。
Fig. 1 Seasonal variations of soil temperature and moisture (mean ± SE).
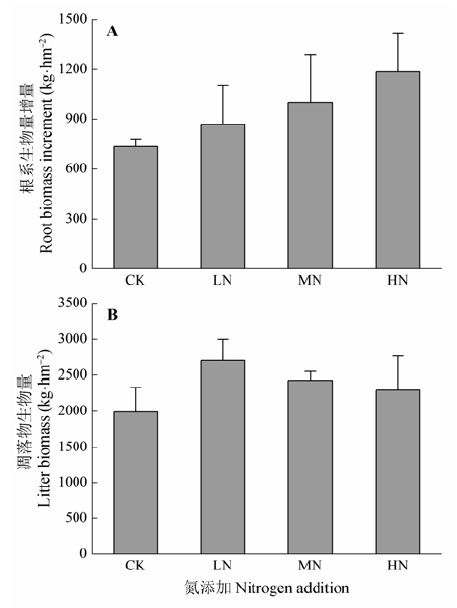
图2 不同氮处理下的根系生物量增量(A)和凋落物生物量(B) (平均值±标准误差)。CK、LN、MN和HN为4种施氮水平, 分别代表氮添加浓度为0、2、5和10 g·m-2·a-1。
Fig. 2 Root biomass increment (A) and litter biomass (B) under different nitrogen treatments (mean ± SE). CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively.
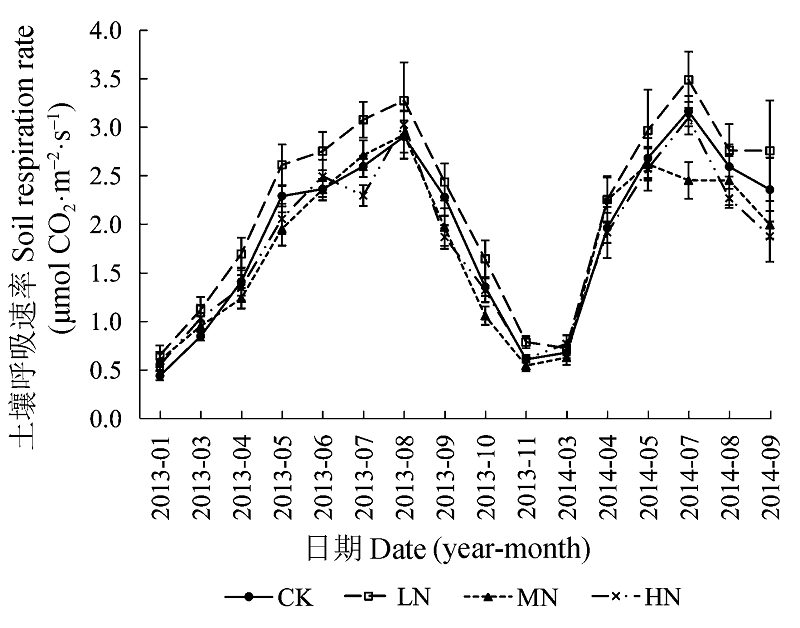
图3 不同氮添加浓度下土壤呼吸的季节动态(平均值±标准误差)。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。
Fig. 3 Seasonal variations of soil respiration under different nitrogen treatments (mean ± SE). CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively.
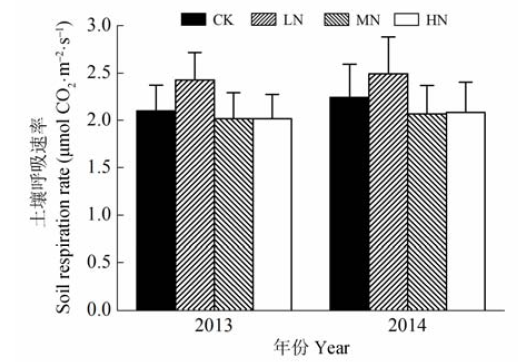
图4 不同氮添加浓度下的生长季土壤呼吸速率(平均值±标准误差)。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。
Fig. 4 Soil respiration rate in the growing season under different nitrogen treatments (mean ± SE). CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively.
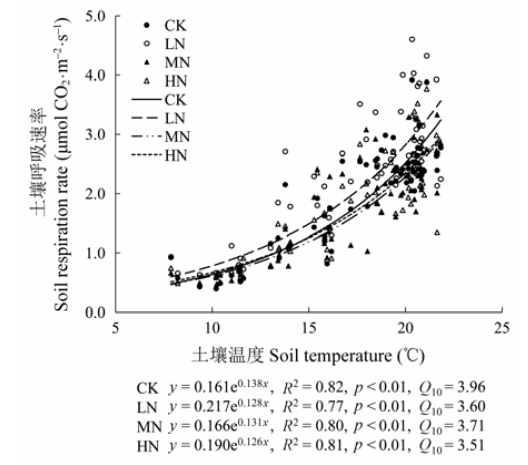
图5 不同氮添加水平下5 cm深度土壤呼吸与土壤温度的关系。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。Q10, 土壤呼吸的温度敏感性。
Fig. 5 Relationships between soil respiration rate and soil temperature at 5 cm soil depth under different nitrogen treatments. CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively. Q10, temperature sensitivity.
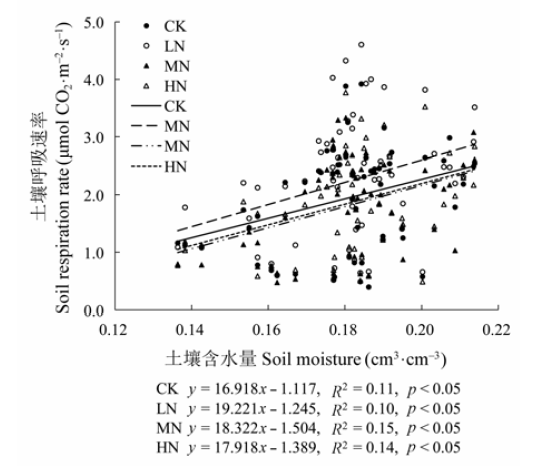
图6 不同氮添加水平下5 cm深度土壤呼吸与土壤含水量的关系。CK、LN、MN和HN为4种施氮水平, 分别代表的氮添加浓度为0、2、5和10 g·m-2·a-1。
Fig. 6 Relationships between soil respiration rate and soil moisture at 5 cm soil depth under different nitrogen treatments. CK, LN, MN and HN stand for 0, 2, 5 and 10 g·m-2·a-1 nitrogen addition, respectively.
| [1] | Aber JD, Nadelhoffer KJ, Steudler P, Melillo JM (1989). Nitrogen saturation in northern forest ecosystems.Bioscience, 39, 378-386. |
| [2] | Bond-Lamberty B, Thomson A (2010). A global database of soil respiration data.Biogeosciences, 7, 1915-1926. |
| [3] | Bowden RD, Davidson E, Savage K, Arabia C, Steudler P (2004). Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest.Forest Ecology and Management, 196, 43-56. |
| [4] | Burton AJ, Pregitzer KS, Crawford JN, Zogg GP, Zak DR (2004). Simulated chronic NO3- deposition reduces soil respiration in northern hardwood forests.Global Change Biology, 10, 1080-1091. |
| [5] | Craine JM, Wedin DA, Reich PB (2001). Grassland species effects on soil CO2 flux track the effects of elevated CO2 and nitrogen.New Phytologist, 150, 425-434. |
| [6] | Cusack DF, Silver WL, Torn MS, McDowell WH (2011). Effects of nitrogen additions on above- and below-ground carbon dynamics in two tropical forests.Biogeochemistry, 104, 203-225. |
| [7] | Davidson EA, Janssens IA (2006). Temperature sensitivity of soil carbon decomposition and feedbacks to climate change.Nature, 440, 165-173. |
| [8] | Denman KL, Brasseur G, Chidthaisong A, Ciais P, Cox PM, Dickinson RE, Hauglustaine D, Heinze C, Holland E, Jacob D, Lohmann U, amachandran S, da Silva Dias PL, Wofsy SC, Zhang X (2007). Couplings between changes in the climate system and biogeochemistry. In: Solomon S, Qin D, Manning M, Marquis M, Averyt K, Tignor MMB, Miller HLR, Chen Z eds. Climate Change 2007, The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. RCambridge University Press, Cambridge, UK. |
| [9] | Deng Q, Zhou G, Liu J, Liu S, Duan H, Zhang D (2010). Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China.Biogeosciences, 7, 315-328. |
| [10] | Dilustro JJ, Collins B, Duncan L, Crawford C (2005). Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests.Forest Ecology and Management, 204, 85-95. |
| [11] | Duan HL, Liu JX, Deng Q, Chen XM, Zhang DQ (2009). Effects of elevated CO2 and N deposition on plant biomass accumulation and allocation in subtropical forest ecosystems: A mesocosm study.Journal of Plant Ecology (Chinese Version), 33, 570-579. (in Chinese with English abstract)[段洪浪, 刘菊秀, 邓琦, 陈小梅, 张德强 (2009). CO2浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响. 植物生态学报, 33, 570-579.] |
| [12] | Egerton-Warburton L, Allen E (2000). Shifts in the diversity of arbuscular mycorrhizal fungi along an anthropogenic nitrogen gradient.Ecological Applications, 10, 484-496. |
| [13] | Elser JJ, Andersen T, Baron JS, Bergstroem A-K, Jansson M, Kyle M, Nydick KR, Steger L, Hessen DO (2009). Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition.Science, 326, 835-837. |
| [14] | Fang H, Mo JM, Peng SL, Li ZA, Wang H (2007). Cumulative effects of nitrogen additions on litter decomposition in three tropical forests in southern China.Plant and Soil, 297, 233-242. |
| [15] | Forster P, Ramaswamy V, Artaxo P, Berntsen T, Betts R, Fahey DW, Haywood J, Lean J, Lowe DC, Myhre G, Nganga J, Prinn R, Raga G, Schulz M, van Dorland R (2007). Changes in atmospheric constituents and in radiative forcing. In: Solomon S, Qin D, Manning M, Marquis M, Averyt K, Tignor MMB, Miller HLR, Chen Z eds. Climate Change 2007, The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. |
| [16] | Frey SD, Knorr M, Parrent JL, Simpson RT (2004). Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests.Forest Ecology and Management, 196, 159-171. |
| [17] | Gallo M, Amonette R, Lauber C, Sinsabaugh RL, Zak DR (2004). Microbial community structure and oxidative enzyme activity in nitrogen-amended north temperate forest soils.Microbial Ecology, 48, 218-229. |
| [18] | Han GX, Zhou GS (2009). Review of spatial and temporal variations of soil respiration and driving mechanisms.Chinese Journal of Plant Ecology, 33, 197-205. (in Chinese with English abstract)[韩广轩, 周广胜 (2009). 土壤呼吸作用时空动态变化及其影响机制研究与展望. 植物生态学报, 33, 197-205.] |
| [19] | Hanson PJ, Edwards NT, Garten CT, Andrews JA (2000). Separating root and soil microbial contributions to soil respiration: A review of methods and observations.Biogeochemistry, 48, 115-146. |
| [20] | Högberg P (2007). Environmental science: Nitrogen impacts on forest carbon.Nature, 447, 781-782. |
| [21] | Högberg P, Fan HB, Quist M, Binkley D, Tamm CO (2006). Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest.Global Change Biology, 12, 489-499. |
| [22] | Hu HF, Wang ZH, GH, Fu BJ (2006). Vegetation carbon storage of major shrublands in China.Journal of Plant Eco- logy (Chinese Version), 30, 539-544. (in Chinese with English abstract)[胡会峰, 王志恒, 刘国华, 傅伯杰 (2006). 中国主要灌丛植被碳储量. 植物生态学报, 30, 539-544.] |
| [23] | Hyvonen R, Agren GI, Linder S, Persson T, Cotrufo MF, Ekblad A, Freeman M, Grelle A, Janssens IA, Jarvis PG, Kellomaki S, Lindroth A, Loustau D, Lundmark T, Norby RJ, Oren R, Pilegaard K, Ryan MG, Sigurdsson BD, Stromgren M, van Oijen M, Wallin G (2007). The likely impact of elevated CO2, nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review.New Phytologist, 173, 463-480. |
| [24] | Hyvonen R, Persson T, Andersson S, Olsson B, Agren GI, Linder S (2008). Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe.Biogeochemistry, 89, 121-137. |
| [25] | IPCC (Intergovernmental Panel on Climate Change) (2013). Contribution of working group 1 to the fifth assessment report of the intergovernmental panel on climate change. In: Stocker TF, Qin DH, Plattner GK, Tignor MMB, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM eds. Climate Change 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK. |
| [26] | Janssens IA, Dieleman W, Luyssaert S, Subke JA, Reichstein M, Ceulemans R, Ciais P, Dolman AJ, Grace J, Matteucci G, Papale D, Piao SL, Schulze ED, Tang J, Law BE (2010). Reduction of forest soil respiration in response to nitrogen deposition.Nature Geoscience, 3, 315-322. |
| [27] | Knorr M, Frey SD, Curtis PS (2005). Nitrogen additions and litter decomposition: A meta-analysis.Ecology, 86, 3252-3257. |
| [28] | LeBauer DS, Treseder KK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.Ecology, 89, 371-379. |
| [29] | Li JX, Zhang X, Xie ZQ, Lu CF, Tu XY, Xun Y (2015). Community composition and structure ofRhododendron simsii shrubland in the Dawei Mountain, Hunan Province. Biodiversity Science, 23, 815-823. (in Chinese with English abstract)[李家湘, 张旭, 谢宗强, 卢从发, 涂向阳, 寻院 (2015). 湖南大围山杜鹃灌丛的群落组成及结构特征. 生物多样性, 23, 815-823.] |
| [30] | Liu X, Duan L, Mo J, Du E, Shen J, Lu X, Zhang Y, Zhou X, He C, Zhang F (2011). Nitrogen deposition and its ecological impact in China: An overview.Environmental Pollution, 159, 2251-2264. |
| [31] | Lue CQ, Tian HQ (2007). Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. Journal of Geophysical Research-Atmospheres, 112, D22S05, doi: 10.1029/2006JD007990. |
| [32] | Luo L, Sheng GZ, Xie ZQ, Zhou LG (2011). Components of soil respiration and its temperature sensitivity in four types of forests along an elevational gradient in Shennongjia, China.Chinese Journal of Plant Ecology, 35, 722-730. (in Chinese with English abstract)[罗璐, 申国珍, 谢宗强, 周利光 (2011). 神农架海拔梯度上4种典型森林的土壤呼吸组分及其对温度的敏感性. 植物生态学报, 35, 722-730.] |
| [33] | Luo YQ, Hui DF, Zhang DQ (2006). Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: A meta-analysis.Ecology, 87, 53-63. |
| [34] | Ma YC, Zhu B, Sun ZZ, Zhao C, Yang Y, Piao SL (2014). The effects of simulated nitrogen deposition on extracellular enzyme activities of litter and soil among different-aged stands of larch.Journal of Plant Ecology, 7, 240-249. |
| [35] | Maskell LC, Smart SM, Bullock JM, Thompson K, Stevens CJ (2010). Nitrogen deposition causes widespread loss of species richness in British habitats.Global Change Biology, 16, 671-679. |
| [36] | Melillo JM, Steudler PA, Aber JD, Newkirk K, Lux H, Bowles FP, Catricala C, Magill A, Ahrens T, Morrisseau S (2002). Soil warming and carbon-cycle feedbacks to the climate system.Science, 298, 2173-2176. |
| [37] | Mo JM, Brown S, Xue JH, Fang YT, Li ZA (2006).Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant and Soil, 282, 135-151. |
| [38] | Mo JM, Zhang W, Zhu WX, Fang YT, Li DJ, Zhao P (2007). Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China.Plant and Soil, 296, 125-135. |
| [39] | Mo JM, Zhang W, Zhu WX, Gundersen P, Fang YT, Li DJ, Wang H (2008). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China.Global Change Biology, 14, 403-412. |
| [40] | Moscatelli MC, Lagornarsino A, de Angelis P, Grego S (2008). Short and mediumterm contrasting effects of nitrogen fertilization on C and N cycling in a poplar plantation soil.Forest Ecology and Management, 255, 447-454. |
| [41] | Pendall E, Bridgham S, Hanson PJ, Hungate B, Kicklighter DW, Johnson DW, Law BE, Luo YQ, Megonigal JP, Olsrud M, Ryan MG, Wan SQ (2004). Belowground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods, and models.New Phytologist, 162, 311-322. |
| [42] | Peng SS, Piao SL, Wang T, Sun JY, Shen ZH (2009). Temperature sensitivity of soil respiration in different ecosystems in China.Soil Biology & Biochemistry, 41, 1008-1014. |
| [43] | Pregitzer KS, Burton AJ, Zak DR, Talhelm AF (2008). Simulated chronic nitrogen deposition increases carbon storage in northern temperate forests.Global Change Biology, 14, 142-153. |
| [44] | Raich JW, Potter CS, Bhagawati D (2002). Interannual varia- bility in global soil respiration, 1980-1994.Global Change Biology, 8, 800-812. |
| [45] | Rodeghiero M, Cescatti A (2006). Indirect partitioning of soil respiration in a series of evergreen forest ecosystems.Plant and Soil, 284, 7-22. |
| [46] | Rühling Å, Tyler G (1991). Effects of simulated nitrogen deposition to the forest floor on the macrofungal flora of a beech forest.Ambio, 20, 261-263. |
| [47] | Ryan MG, Law BE (2005). Interpreting, measuring, and modeling soil respiration.Biogeochemistry, 73, 3-27. |
| [48] | Schlesinger WH, Andrews JA (2000). Soil respiration and the global carbon cycle.Biogeochemistry, 48, 7-20. |
| [49] | Sotta ED, Meir P, Malhi Y, Nobre AD, Hodnett M, Grace J (2004). Soil CO2 efflux in a tropical forest in the central Amazon.Global Change Biology, 10, 601-617. |
| [50] | Sun ZZ, Liu LL, Ma YC, Yin GD, Zhao C, Zhang Y, Piao SL (2014). The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil.Agricultural and Forest Meteorology, 197, 103-110. |
| [51] | Tang XL, Liu SG, Zhou GY, Zhang DQ, Zhou CY (2006). Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China.Global Change Biology, 12, 546-560. |
| [52] | Tarnocai C, Canadell JG, Schuur EAG, Kuhry P, Mazhitova G, Zimov S (2009). Soil organic carbon pools in the northern circumpolar permafrost region. Global Biogeochemical Cycles, 23, GB2023, doi: 10.1029/2008GB003327. |
| [53] | Thomas RQ, Canham CD, Weathers KC, Goodale CL (2010). Increased tree carbon storage in response to nitrogen deposition in the US.Nature Geoscience, 3, 13-17. |
| [54] | Tu LH, Hu TX, Zhang J, Li XW, Hu HL, Liu L, Xiao YL (2013). Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem.Soil Biology & Biochemistry, 58, 255-264. |
| [55] | Xu WH, Wan SQ (2008). Water- and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China.Soil Biology & Biochemistry, 40, 679-687. |
| [56] | Zheng SW, Tang M, Zou JH, Mu CL (2007). Summary of research on shrub biomass in China.Journal of Chengdu University (Natural Science Edition), 26, 189-192. (in Chinese with English abstract)[郑绍伟, 唐敏, 邹俊辉, 慕长龙 (2007). 灌木群落及生物量研究综述. 成都大学学报(自然科学版), 26, 189-192.] |
| [57] | Zheng XH, Fu CB, Xu XK, Yan XD, Huang Y, Han SH, Hu F, Chen GX (2002). The Asian nitrogen cycle case study.AMBIO, 31, 79-87. |
| [58] | Zheng ZM, Yu GR, Fu YL, Wang YS, Sun XM, Wang YH (2009). Temperature sensitivity of soil respiration is affected by prevailing climatic conditions and soil organic carbon content: A trans-China based case study.Soil Biology & Biochemistry, 41, 1531-1540. |
| [59] | Zhou LY, Zhou XH, Zhang BC, Lu M, Luo YQ, Liu LL, Li B (2014). Different responses of soil respiration and its components to nitrogen addition among biomes: A meta- analysis.Global Change Biology, 20, 2332-2343. |
| [1] | 沈健, 何宗明, 董强, 郜士垒, 林宇. 轻度火烧对滨海沙地人工林土壤呼吸速率和非生物因子的影响[J]. 植物生态学报, 2023, 47(7): 1032-1042. |
| [2] | 李雪, 董杰, 韩广轩, 张奇奇, 谢宝华, 李培广, 赵明亮, 陈克龙, 宋维民. 黄河三角洲典型滨海盐沼湿地土壤CO2和CH4排放对水盐变化的响应[J]. 植物生态学报, 2023, 47(3): 434-446. |
| [3] | 魏春雪, 杨璐, 汪金松, 杨家明, 史嘉炜, 田大栓, 周青平, 牛书丽. 实验增温对陆地生态系统根系生物量的影响[J]. 植物生态学报, 2021, 45(11): 1203-1212. |
| [4] | 李建军, 刘恋, 陈迪马, 许丰伟, 程军回, 白永飞. 底座入土深度和面积对典型草原土壤呼吸测定结果的影响[J]. 植物生态学报, 2019, 43(2): 152-164. |
| [5] | 王祥, 朱亚琼, 郑伟, 关正翾, 盛建东. 昭苏山地草甸4种典型土地利用方式下的土壤呼吸特征[J]. 植物生态学报, 2018, 42(3): 382-396. |
| [6] | 岑宇, 王成栋, 张震, 任侠, 刘美珍, 杨帆. 河北省天然草地生物量和碳密度空间分布格局[J]. 植物生态学报, 2018, 42(3): 265-276. |
| [7] | 柴曦, 李英年, 段呈, 张涛, 宗宁, 石培礼, 何永涛, 张宪洲. 青藏高原高寒灌丛草甸和草原化草甸CO2通量动态及其限制因子[J]. 植物生态学报, 2018, 42(1): 6-19. |
| [8] | 朱志成, 黄银, 许丰伟, 邢稳, 郑淑霞, 白永飞. 降雨强度和时间频次对内蒙古典型草原土壤氮矿化的影响[J]. 植物生态学报, 2017, 41(9): 938-952. |
| [9] | 葛晓改, 周本智, 肖文发, 王小明, 曹永慧, 叶明. 生物质炭添加对毛竹林土壤呼吸动态和温度敏感性的影响[J]. 植物生态学报, 2017, 41(11): 1177-1189. |
| [10] | 姚辉, 胡雪洋, 朱江玲, 朱剑霄, 吉成均, 方精云. 北京东灵山3种温带森林土壤呼吸及其20年的变化[J]. 植物生态学报, 2015, 39(9): 849-856. |
| [11] | 许洺山, 黄海侠, 史青茹, 杨晓东, 周刘丽, 赵延涛, 张晴晴, 阎恩荣. 浙东常绿阔叶林植物功能性状对土壤含水量变化的响应[J]. 植物生态学报, 2015, 39(9): 857-866. |
| [12] | 王铭, 刘兴土, 张继涛, 李秀军, 王国栋, 鲁新蕊, 李晓宇. 松嫩平原西部草甸草原5种典型植物群落土壤呼吸的时空动态[J]. 植物生态学报, 2014, 38(4): 396-404. |
| [13] | 李悦, 刘颖慧, 申卫军, 徐霞, 田玉强. 内蒙古克氏针茅草原土壤异养呼吸对土壤温度和水分变化的响应[J]. 植物生态学报, 2014, 38(3): 238-248. |
| [14] | 吴君君, 杨智杰, 刘小飞, 熊德成, 林伟盛, 陈朝琪, 王小红. 米槠和杉木人工林土壤呼吸及其组分分析[J]. 植物生态学报, 2014, 38(1): 45-53. |
| [15] | 邸楠,席本野,Jeremiah R.PINTO,王烨,李广德,贾黎明. 宽窄行栽植下三倍体毛白杨根系生物量分布及其对土壤养分因子的响应[J]. 植物生态学报, 2013, 37(10): 961-971. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19