
植物生态学报 ›› 2018, Vol. 42 ›› Issue (9): 926-937.DOI: 10.17521/cjpe.2018.0167
沈芳芳1,2,李燕燕1,2,刘文飞2,段洪浪2,樊后保2,*( ),胡良2,孟庆银3
),胡良2,孟庆银3
收稿日期:2018-07-16
修回日期:2018-09-10
出版日期:2018-09-20
发布日期:2019-01-15
通讯作者:
樊后保
基金资助:
SHEN Fang-Fang1,2,LI Yan-Yan1,2,LIU Wen-Fei2,DUAN Hong-Lang2,FAN Hou-Bao2,*(),HU Liang2,MENG Qing-Yin3
Received:2018-07-16
Revised:2018-09-10
Online:2018-09-20
Published:2019-01-15
Contact:
Hou-Bao FAN
Supported by:摘要:
为了解森林养分内循环对全球变化的响应, 基于长期模拟氮沉降试验, 研究了杉木(Cunninghamia lanceolata)人工林不同龄级(一年生、二年生和衰老)叶和枝的氮(N)、磷(P)养分分配及其再吸收特征, 并分析了不同模拟N沉降处理时间(7年和14年)杉木叶N、P养分再吸收差异。在12年生杉木中开展模拟N沉降试验, 以尿素(CO(NH2)2)为N源, 设N0、N1、N2和N3 4个处理水平, 施氮量分别为0、60、120和240 kg·hm -2·a -1, 每个处理重复3次。结果表明: (1)叶和枝在衰老过程中碳(C)、N和P含量逐渐降低, 且叶的C、N和P含量比枝高; N含量大小依次为一年生叶>二年生叶>衰老叶>一年生枝>二年生枝>衰老枝, 且N3 > N2 > N1 > N0, 而C:N则呈现相反的趋势; 衰老器官的C:N、C:P、N:P比新鲜器官高; N沉降增加了不同龄级叶和枝(除二年生叶外)的N、N:P和C:P, 但降低了P和C:N。(2)叶和枝的N、P养分再吸收率(REN、REP)随龄级的增加至衰老有规律地递减, 且REP > REN; 受长期N沉降的影响, REN叶(28.12%) <枝(30.00%), 而REP则为叶(45.82%) >枝(30.42%); 杉木叶和枝N:P与REN:REP之间存在极显著的线性相关关系。(3)随N沉降处理时间的增加, 叶REN呈降低态势, 各处理(N1、N2和N3)分别降低了9.85%、3.17%和11.71%; 而REP则明显上升, 分别增加了71.98%、42.25%和9.60%。研究结果表明: 不同器官、不同龄级的养分再吸收率随氮沉降处理的水平、处理时间而所有不同; REN:REP与N:P之间存在紧密关系。
沈芳芳, 李燕燕, 刘文飞, 段洪浪, 樊后保, 胡良, 孟庆银. 长期氮沉降对杉木人工林叶、枝氮磷养分再吸收的影响. 植物生态学报, 2018, 42(9): 926-937. DOI: 10.17521/cjpe.2018.0167
SHEN Fang-Fang, LI Yan-Yan, LIU Wen-Fei, DUAN Hong-Lang, FAN Hou-Bao, HU Liang, MENG Qing-Yin. Responses of nitrogen and phosphorus resorption from leaves and branches to long-term nitrogen deposition in a Chinese fir plantation. Chinese Journal of Plant Ecology, 2018, 42(9): 926-937. DOI: 10.17521/cjpe.2018.0167
| 处理 Treatment | 林分特征 Stand characteristics | 土壤理化性质 Soil physicochemical properties | ||||||
|---|---|---|---|---|---|---|---|---|
| 林龄 Forest age (a) | 密度 Density (No.·hm-2) | 平均胸径 Mean DBH (cm) | 平均树高 Mean tree height (m) | pH | 有机碳 Organic carbon (g·kg-1) | 全氮 Total N (g·kg-1) | 全磷 Total P (g·kg-1) | |
| N0 | 12 | 1 717 | 16.1 | 11.8 | 4.59 | 19.23 | 0.86 | 0.22 |
| N1 | 12 | 1 633 | 16.0 | 12.2 | 4.76 | 17.31 | 0.68 | 0.13 |
| N2 | 12 | 1 683 | 16.3 | 12.2 | 4.65 | 18.88 | 0.80 | 0.17 |
| N3 | 12 | 1 625 | 16.0 | 12.1 | 4.71 | 18.14 | 0.81 | 0.14 |
表1 各处理样地的主要林分特征和土壤理化性质本底值
Table 1 Background values of the stand and soil physicochemical properties in the nitrogen addition plots
| 处理 Treatment | 林分特征 Stand characteristics | 土壤理化性质 Soil physicochemical properties | ||||||
|---|---|---|---|---|---|---|---|---|
| 林龄 Forest age (a) | 密度 Density (No.·hm-2) | 平均胸径 Mean DBH (cm) | 平均树高 Mean tree height (m) | pH | 有机碳 Organic carbon (g·kg-1) | 全氮 Total N (g·kg-1) | 全磷 Total P (g·kg-1) | |
| N0 | 12 | 1 717 | 16.1 | 11.8 | 4.59 | 19.23 | 0.86 | 0.22 |
| N1 | 12 | 1 633 | 16.0 | 12.2 | 4.76 | 17.31 | 0.68 | 0.13 |
| N2 | 12 | 1 683 | 16.3 | 12.2 | 4.65 | 18.88 | 0.80 | 0.17 |
| N3 | 12 | 1 625 | 16.0 | 12.1 | 4.71 | 18.14 | 0.81 | 0.14 |
| 因子 Factor | F (p)值 F (p) value | |||||
|---|---|---|---|---|---|---|
| C | N | P | C:N | C:P | N:P | |
| 叶组分 Leaf components | 1.173 (0.336) | 20.401 (<0.001) | 24.425 (<0.001) | 13.832 (<0.001) | 24.072 (<0.001) | 5.728 (0.003) |
| 氮沉降 N deposition | 0.573 (0.637) | 3.272 (0.034) | 1.246 (0.309) | 3.158 (0.038) | 0.853 (0.475) | 0.631 (0.601) |
| 叶组分×氮沉降 Leaf components × N deposition | 0.280 (0.976) | 0.893 (0.543) | 0.865 (0.565) | 0.537 (0.837) | 0.963 (0.487) | 0.575 (0.807) |
| 枝组分 Branch components | 1.191 (0.329) | 9.407 (<0.001) | 19.685 (<0.001) | 6.534 (0.001) | 6.010 (0.002) | 8.176 (<0.001) |
| 氮沉降 N deposition | 0.165 (0.919) | 1.782 (0.170) | 2.130 (0.116) | 1.229 (0.315) | 1.063 (0.378) | 5.123 (0.005) |
| 枝组分×氮沉降 Branch components × N deposition | 0.398 (0.927) | 0.608 (0.781) | 0.365 (0.943) | 0.429 (0.910) | 0.371 (0.940) | 0.442 (0.902) |
表2 基于双因素方差分析得到的组分、氮沉降水平及其交互作用对杉木叶和枝的生态化学计量的影响(F值)
Table 2 Results (F values) of two-way ANOVA on the effects of components, nitrogen deposition treatment level and their interaction the ecological stoichiometry in leaves and branches of Chinese fir
| 因子 Factor | F (p)值 F (p) value | |||||
|---|---|---|---|---|---|---|
| C | N | P | C:N | C:P | N:P | |
| 叶组分 Leaf components | 1.173 (0.336) | 20.401 (<0.001) | 24.425 (<0.001) | 13.832 (<0.001) | 24.072 (<0.001) | 5.728 (0.003) |
| 氮沉降 N deposition | 0.573 (0.637) | 3.272 (0.034) | 1.246 (0.309) | 3.158 (0.038) | 0.853 (0.475) | 0.631 (0.601) |
| 叶组分×氮沉降 Leaf components × N deposition | 0.280 (0.976) | 0.893 (0.543) | 0.865 (0.565) | 0.537 (0.837) | 0.963 (0.487) | 0.575 (0.807) |
| 枝组分 Branch components | 1.191 (0.329) | 9.407 (<0.001) | 19.685 (<0.001) | 6.534 (0.001) | 6.010 (0.002) | 8.176 (<0.001) |
| 氮沉降 N deposition | 0.165 (0.919) | 1.782 (0.170) | 2.130 (0.116) | 1.229 (0.315) | 1.063 (0.378) | 5.123 (0.005) |
| 枝组分×氮沉降 Branch components × N deposition | 0.398 (0.927) | 0.608 (0.781) | 0.365 (0.943) | 0.429 (0.910) | 0.371 (0.940) | 0.442 (0.902) |
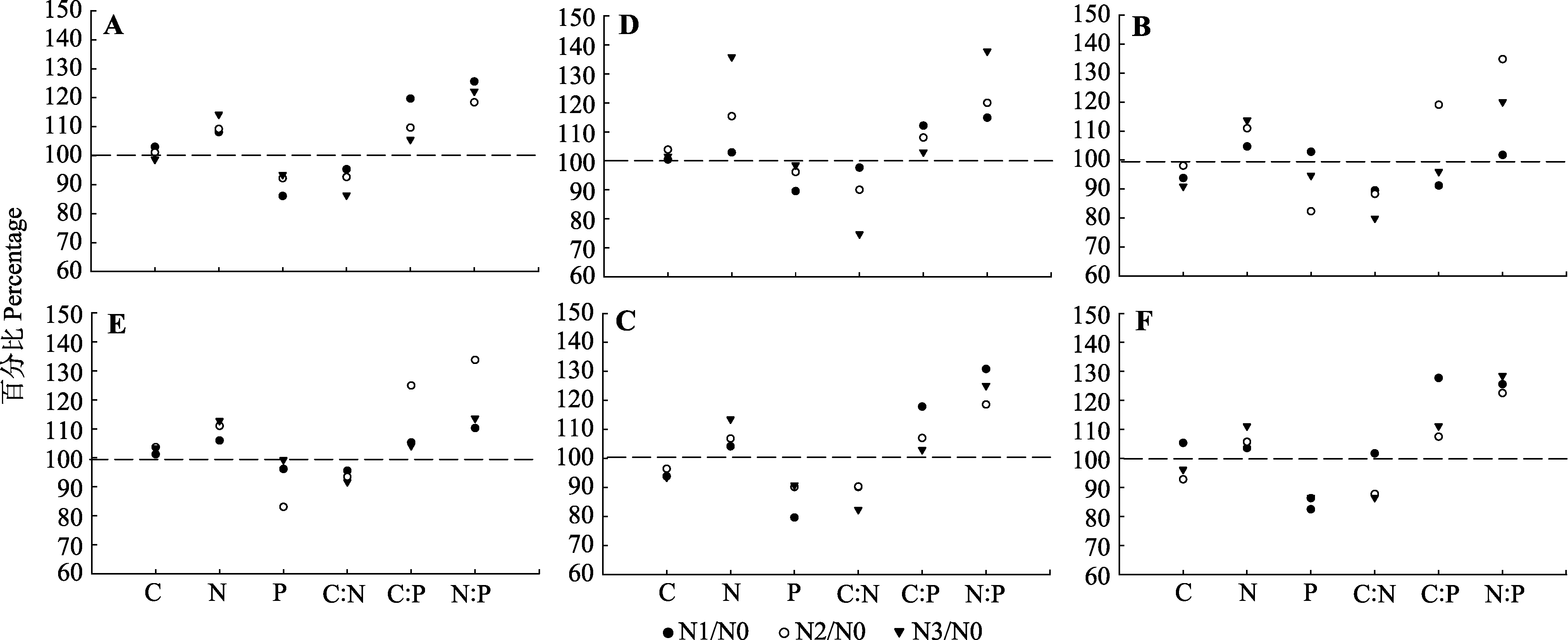
图2 杉木叶和枝的测试指标(C、N、P、C:N、C:P、N:P)氮沉降处理平均值与N0处理平均值的百分比(N0/N0 = 100%)。A, 一年生叶。B, 二年生叶。C, 衰老叶。D, 一年生枝。E, 二年生枝。F, 衰老枝。N0、N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 2 Mean percentage changes of measured parameters (C, N, P, C:N, C:P, N:P) of Chinese fir leaf and branch after N deposition treatment for 14 years relative to these values under control (N0) (N0/N0 = 100%). A, One-year old leaf. B, Two-year old leaf. C, Senesced leaf. D, One-year old branch. E, Two-year old branch. F, Senesced branch. N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
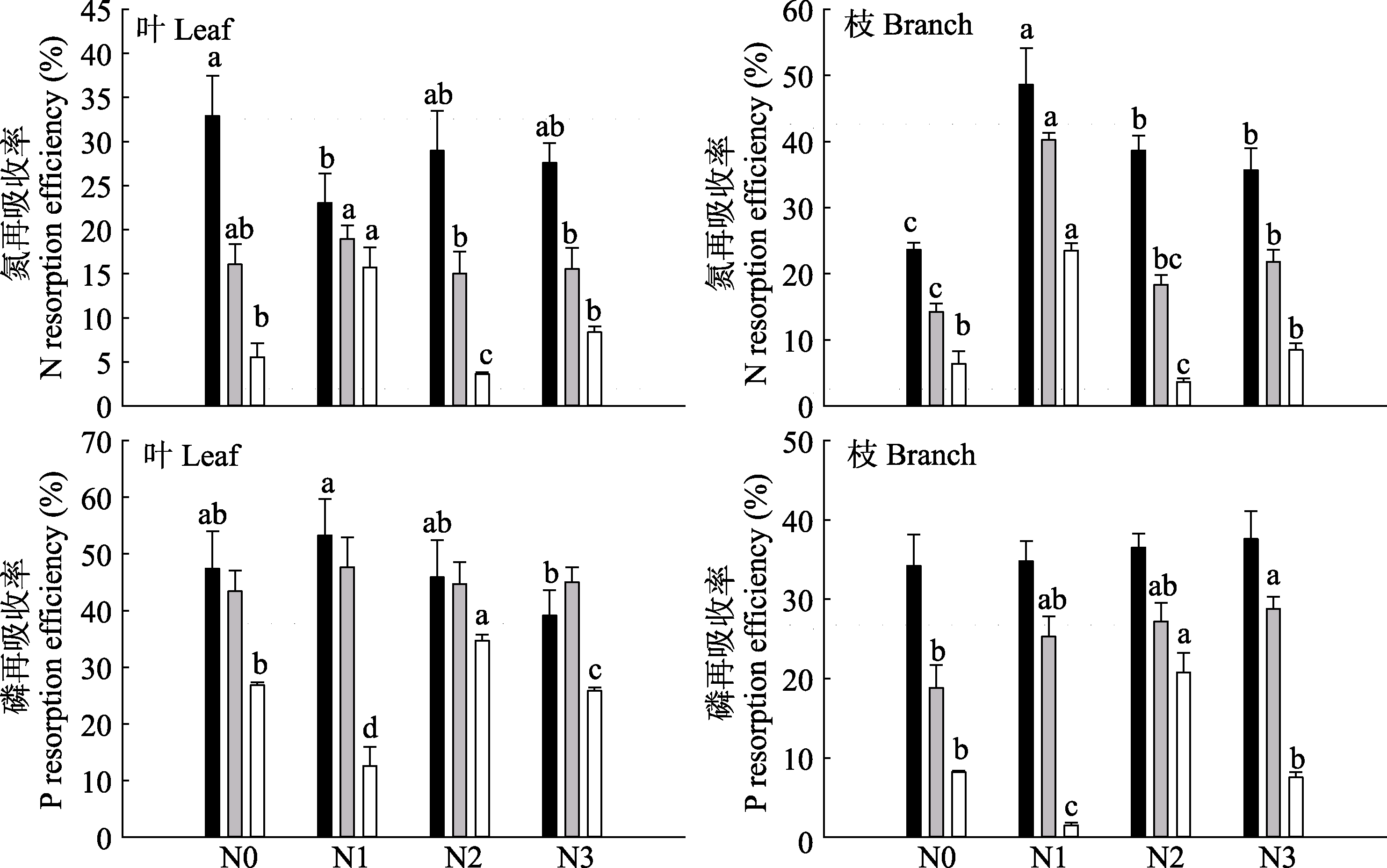
图3 长期氮沉降条件下杉木叶和枝的N、P再吸收率(平均值+标准误差)。黑色柱子表示一年生叶/枝与衰老叶/枝之间的养分再吸收率; 浅灰柱子表示二年生叶/枝与衰老叶/枝之间的养分再吸收率; 白色柱子表示一年生叶/枝与二年生叶/枝之间的养分再吸收率。N0、N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。不同小写字母表示不同氮沉降水平下达到显著差异水平(p < 0.05)。
Fig. 3 Effects of long-term nitrogen deposition on N and P resorption efficiency in leaves and branches of Chinese fir (mean + SE). The black column indicates the ratio of the nutrient resorption efficiency between one-year old leaves/branches and senescent leaves/branches; light gray column indicates the ratio of the nutrient resorption efficiency between two-year old leaves/branches and senescent leaves/branches; white column indicates the ratio of the nutrient resorption efficiency between one-year old leaves/branches and two-year-old leaves. N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively. Different lowercase letters indicate significant differences under different nitrogen addition treatment levels (p < 0.05).
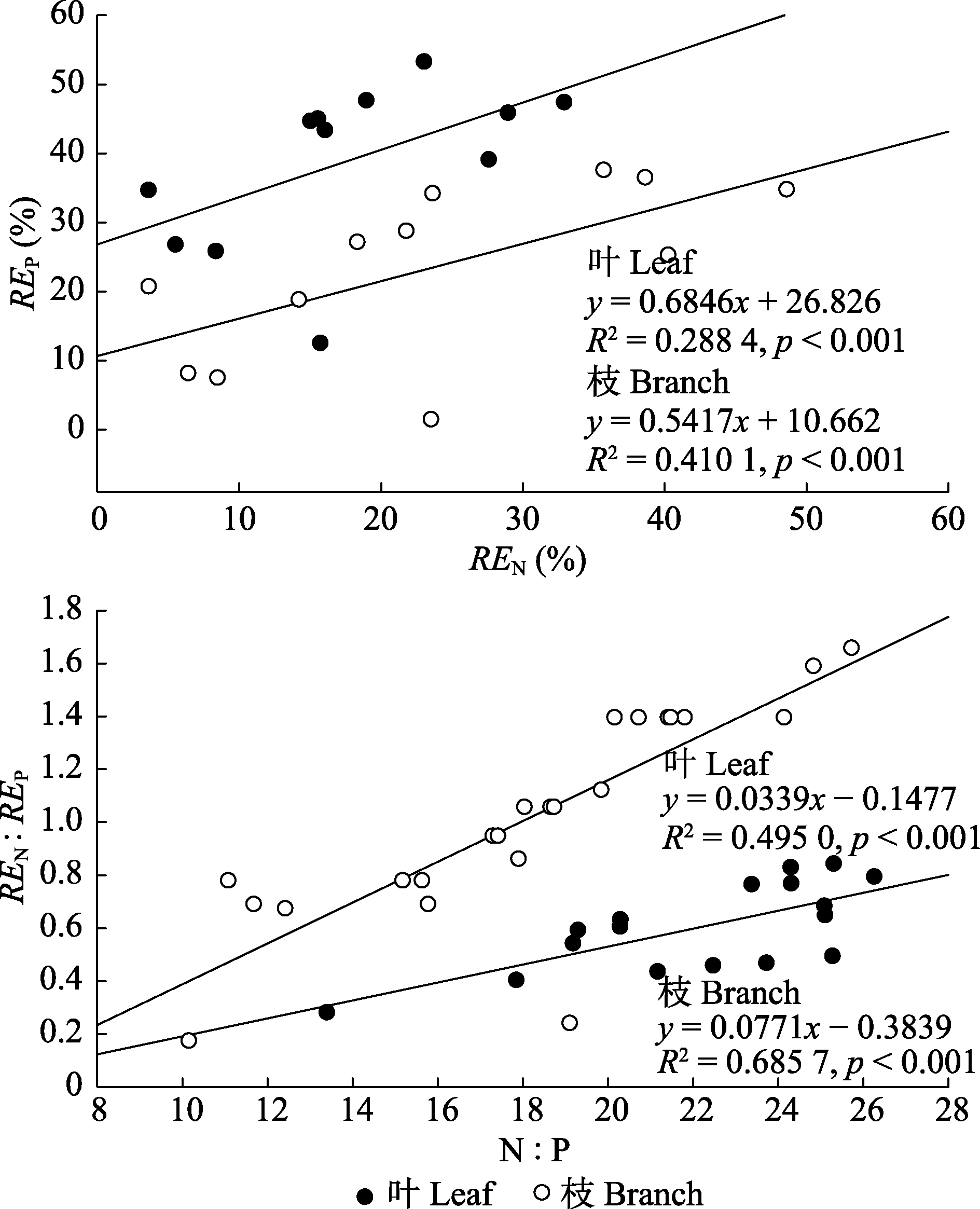
图4 长期氮沉降条件下杉木叶和枝REN与REP, N:P与REN:REP的线性关系。
Fig. 4 Linear relationship between N resorption efficiency (REN) and P resorption efficiency (REP), N:P and N resorption efficiency to P resorption efficiency ratio (REN:REP) of leaf and branch under long-term nitrogen deposition treatments. Data included treatments for 14 years.
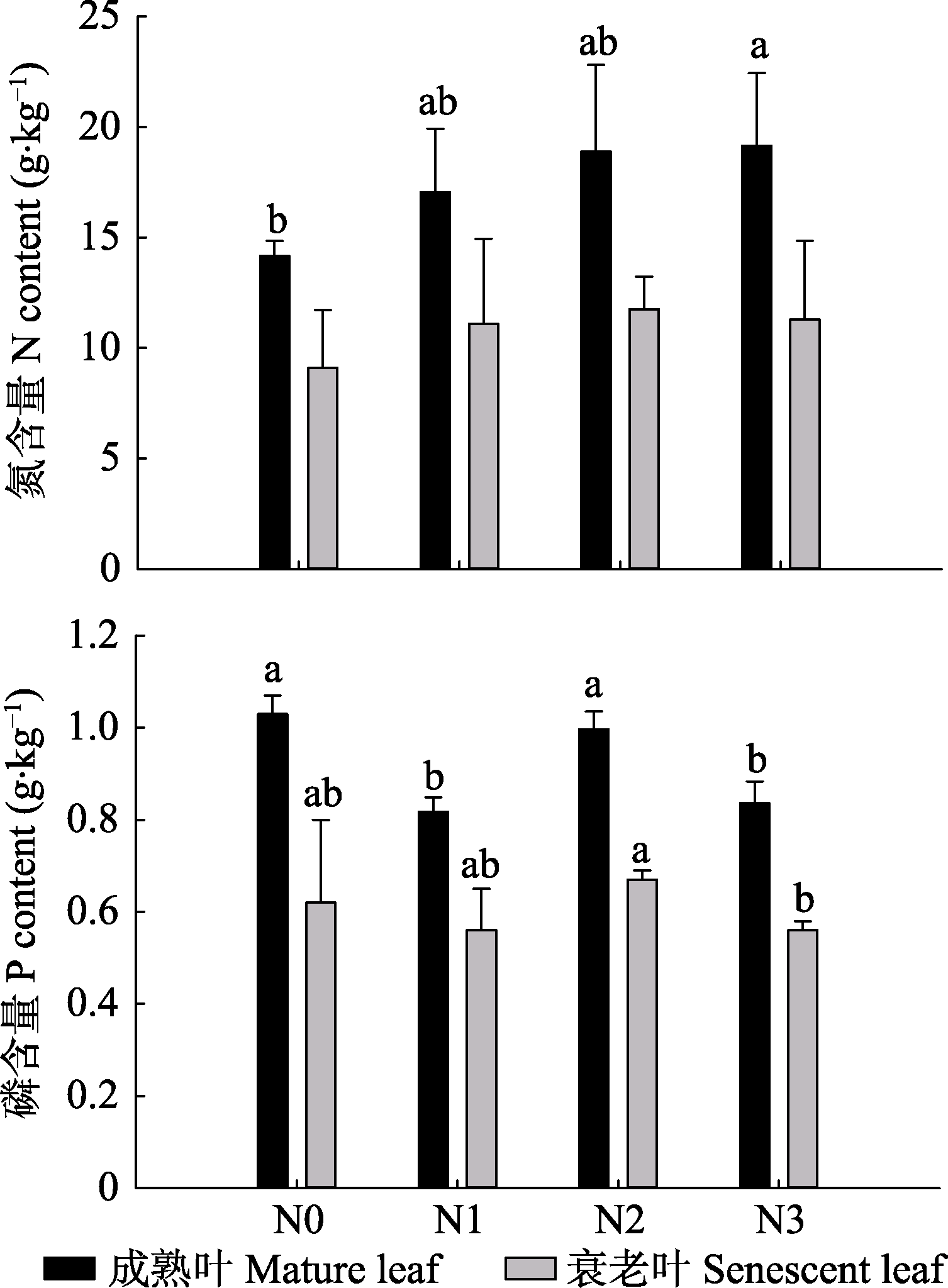
图5 2010年成熟绿叶和衰老叶的N、P含量(平均值+标准误差)。N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 5 N and P content of mature living and senescent leaves in 2010 (mean + SE). N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
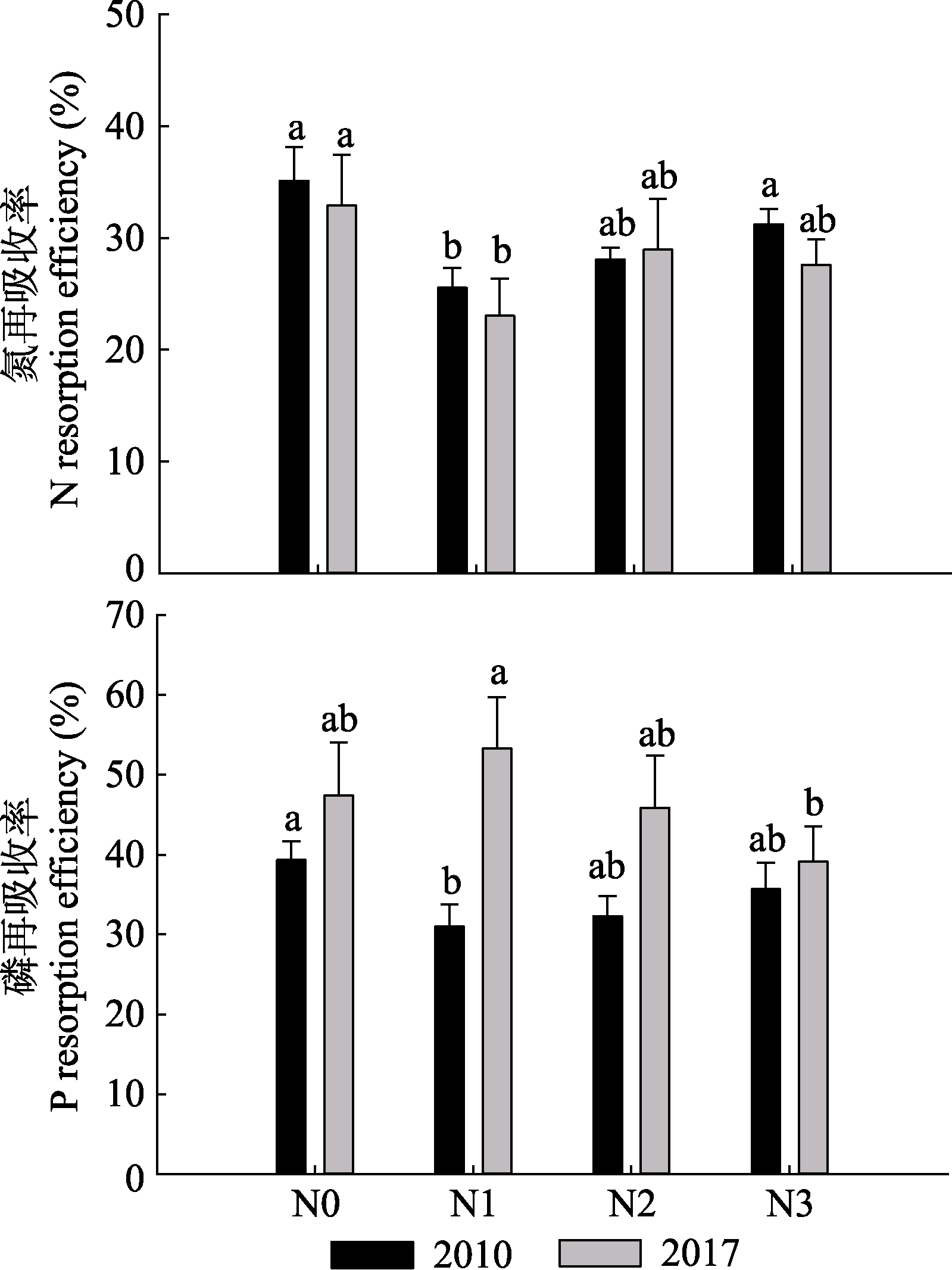
图6 2010和2017年的叶N、P再吸收率(平均值+标准误差)。N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 6 N and P resorption efficiency of leaves in 2010 and 2017 (mean + SE). N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
| 处理 Treatment | 2010 | 2017 | |||
|---|---|---|---|---|---|
| 成熟-衰叶Mature- senescent leaf | 一-衰叶 One-senesced leaf | 二-衰叶 Two-senesced leaf | 一-衰枝 One-senesced branch | 二-衰枝 Two-senesced branch | |
| N0 | 0.89 | 0.69 | 0.37 | 0.69 | 0.75 |
| N1 | 0.82 | 0.43 | 0.40 | 1.40 | 1.59 |
| N2 | 0.87 | 0.63 | 0.34 | 1.06 | 0.67 |
| N3 | 0.88 | 0.70 | 0.35 | 0.95 | 0.76 |
表3 2010和2017年的叶和枝N再吸收率与P再吸收率的比值
Table 3 The ratio of N resorption efficiency to P resorption efficiency (REN:REP) of leaves and branches in 2010 and 2017
| 处理 Treatment | 2010 | 2017 | |||
|---|---|---|---|---|---|
| 成熟-衰叶Mature- senescent leaf | 一-衰叶 One-senesced leaf | 二-衰叶 Two-senesced leaf | 一-衰枝 One-senesced branch | 二-衰枝 Two-senesced branch | |
| N0 | 0.89 | 0.69 | 0.37 | 0.69 | 0.75 |
| N1 | 0.82 | 0.43 | 0.40 | 1.40 | 1.59 |
| N2 | 0.87 | 0.63 | 0.34 | 1.06 | 0.67 |
| N3 | 0.88 | 0.70 | 0.35 | 0.95 | 0.76 |
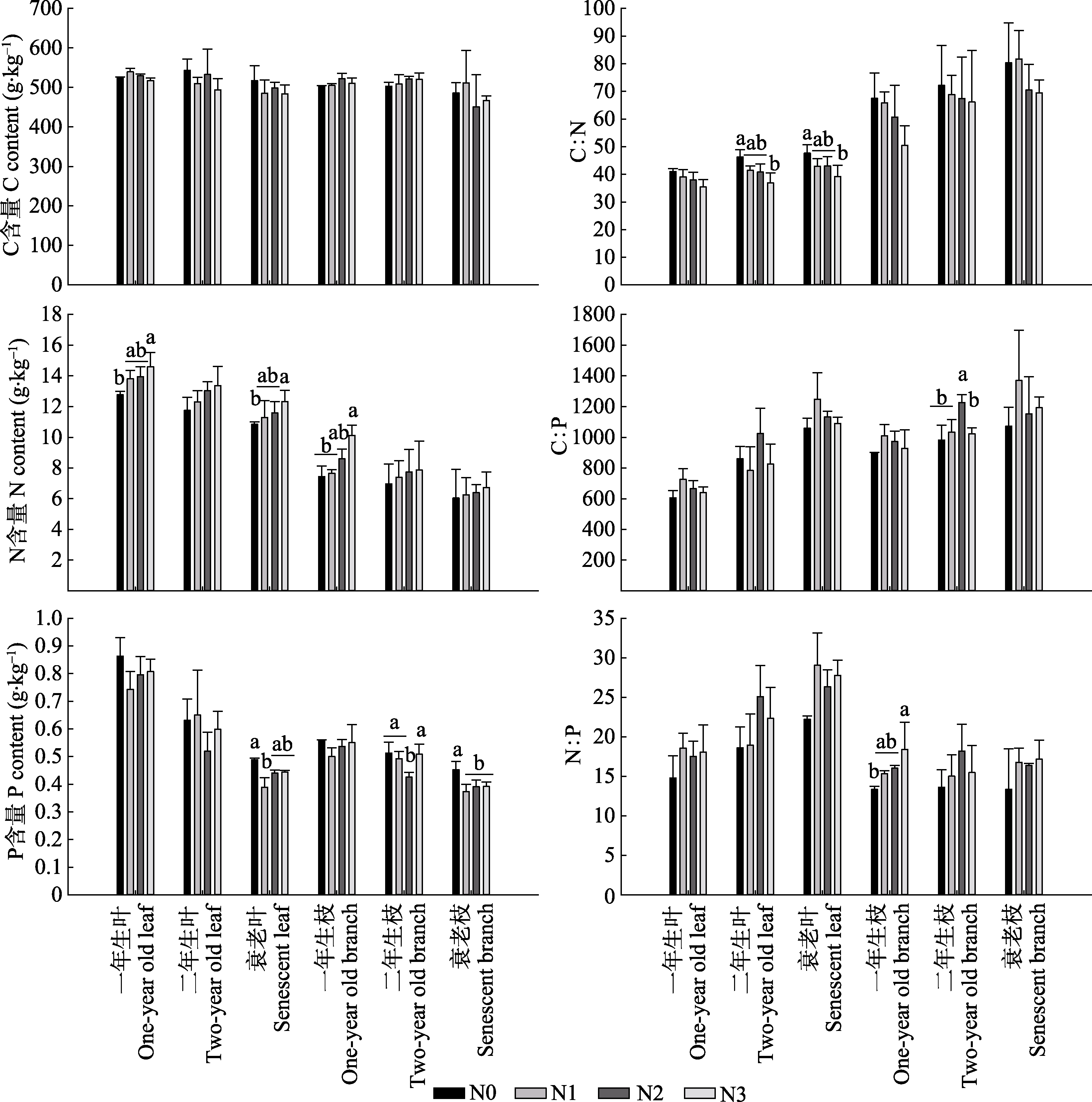
图1 长期氮沉降条件下杉木叶和枝的C、N、P含量及化学计量(平均值+标准误差)。不同小写字母表示相同龄级叶或枝不同氮沉降水平下差异显著(p < 0.05)。N0、N1、N2、N3, 施氮量分别为0、60、120、240 kg·hm-2·a-1。
Fig. 1 C, N, P content and stoichiometry of leaves and branches of Chinese fir under long-term nitrogen deposition (mean + SE). Different lowercase letters indicate significant differences for the same life span leaf or branch under different nitrogen deposition treatment levels (p < 0.05). N0, N1, N2, N3, nitrogen addition 0, 60, 120, 240 kg·hm-2·a-1, respectively.
| [1] |
Aerts R ( 1996). Nutrient resorption from senescing leaves of perennials: Are there general patterns? Journal of Ecology, 84, 597-608.
DOI URL |
| [2] |
Brant AN, Chen HYH ( 2015). Patterns and mechanisms of nutrient resorption in plants. Critical Reviews in Plant Sciences, 34, 471-486.
DOI URL |
| [3] |
Chen FS, Niklas KJ, Chen GS, Guo D ( 2012). Leaf traits and relationships differ with season as well as among species groupings in a managed Southeastern China forest landscape. Plant Ecology, 213, 1489-1502.
DOI URL |
| [4] |
Chen FS, Niklas KJ, Liu Y, Fang XM, Wan SZ, Wang HM ( 2015). Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree physiology, 35, 1106-1117.
DOI URL PMID |
| [5] |
Chen S, Chen SL, Guo ZW ( 2015). Effects of mulching man agement on the internal cycling of nutrients in the rhizomatous roots of Phyllostachys violascens. Acta Ecologica Sinca, 35, 5788-5796.
DOI URL |
|
[ 陈珊, 陈双林, 郭子武 ( 2015). 林地覆盖经营对雷竹鞭根主要养分内循环的影响. 生态学报, 35, 5788-5796.]
DOI URL |
|
| [6] |
Deng Q, Hui DF, Dennis S, Reddy KC ( 2017). Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta-analysis. Global Ecology and Biogeography, 26, 713-728.
DOI URL |
| [7] |
Drenovsky RE, Richards JH ( 2006). Low leaf N and P resorption contributes to nutrient limitation in two desert shrubs. Plant Ecology, 183, 305-314.
DOI URL |
| [8] |
Escudero A, Mediavilla S ( 2003). Decline in photosynthetic nitrogen use efficiency with leaf age and nitrogen resorption as determinants of leaf life span. Journal of Ecology, 91, 880-889.
DOI URL |
| [9] | Fan HB, Liu WF, Li YY, Liao YC, Yuan YH, Xu L ( 2007). Tree growth and soil nutrients in response to nitrogen deposition in a subtropical Chinese fir plantation. Acta Ecologica Sinica, 27, 4630-4642. |
| [ 樊后保, 刘文飞, 李燕燕, 廖迎春, 袁颖红, 徐雷 ( 2007). 亚热带杉木(Cunninghamia lanceolata)人工林生长与土壤养分对氮沉降的响应. 生态学报, 27, 4630-4642.] | |
| [10] |
Fan HB, Su BQ, Lin DX, Chen SP ( 2000). Biogechemical cycle within ecosystem of Chinese fir plantations II: Dynamics of nutrients returning to ecosystem. Chinese Journal of Applied & Environmental Biology, 6, 133-137.
DOI URL |
|
[ 樊后保, 苏兵强, 林德喜, 陈世品 ( 2000). 杉木人工林生态系统的生物地球化学循环II: 氮素沉降动态. 应用与环境生物学报, 6, 133-137].
DOI URL |
|
| [11] |
Fei SM ( 2001). A study on the internal transfer and cycling of nutrients in loblolly pine plantation. Scientia Silvae Sinicae, 37(3), 14-19.
DOI URL |
|
[ 费世民 ( 2001). 火炬松人工林养分体内转移与内循环研究. 林业科学, 37(3), 14-19.]
DOI URL |
|
| [12] |
Fenn ME, Poth MA, Aber JD, Baron JS, Bormann BT, Johnson DW, Lemly AD, McNulty SG, Ryan DF, Stottlemyer R ( 1998). Nitrogen excess in North American ecosystems: Predisposing factors, ecosystem response, and management strategies. Ecological Applications, 8, 706-733.
DOI URL |
| [13] |
Fife D, Nambiar E, Saur E ( 2008). Retranslocation of foliar nutrients in evergreen tree species planted in a Mediterranean environment. Tree Physiology, 28, 187-196.
DOI URL PMID |
| [14] |
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai ZC, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA ( 2008). Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science, 320, 889-892.
DOI URL PMID |
| [15] |
Gundersen P, Emmett BA, Kj?nass OJ, Koopmans CJ, Tietema A ( 1998). Impact of nitrogen deposition on nitrogen cycling in forest: A synthesis of NITREX data. Forest Ecology and Management, 101, 37-55.
DOI URL |
| [16] |
H?egberg P, Fan HB, Quist M, Binkley D, Tamm CO ( 2006). Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest. Global Change Biology, 12, 489-499.
DOI URL |
| [17] |
Huang G, Su YG, Mu XH, Li Y ( 2018). Foliar nutrient resorption responses of three life-form plants to water and nitrogen additions in a temperate desert. Plant and Soil, 424, 479-489.
DOI URL |
| [18] |
Huang JY, Zhu XG, Yuan ZY, Song SH, Li X, Li LH ( 2008). Changes in nitrogen resorption traits of six temperate grassland species along a multi-level N addition gradient. Plant and Soil, 306, 149-158.
DOI URL |
| [19] |
Huang ZQ, He ZM, Wan XH, Hu ZH, Fan SH, Yang YS ( 2013). Harvest residue management effects on tree growth and ecosystem carbon in a Chinese fir plantation in subtropical China. Plant and Soil, 364, 303-314.
DOI URL |
| [20] |
Jiang DL, Xu X, Ruan HH ( 2017). Review of nutrient resorption and its regulating in plants. Journal of Nanjing Forestry University (Natural Sciencies Edition), 41, 183-188.
DOI URL |
|
[ 江大龙, 徐侠, 阮宏华 ( 2017). 植物养分重吸收及其影响研究进展. 南京林业大学学报(自然科学版), 41, 183-188.]
DOI URL |
|
| [21] |
Killingbeck KT ( 1996). Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology, 77, 1716-1727.
DOI URL |
| [22] |
Kobe RK, Lepczyk CA, Iyer M ( 2005). Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology, 86, 2780-2792.
DOI URL |
| [23] |
Li DJ, Mo JM, Fang YT, Cai XA, Xue JH, Xu GL ( 2004). Effects of simulated nitrogen deposition on growth and photosynthesis of Schima superba, Castanopsis chinensis and Cryptocarya concinna seedlings. Acta Ecologica Sinica, 24, 876-882.
DOI URL |
|
[ 李德军, 莫江明, 方运霆, 蔡锡安, 薛璟花, 徐国良 ( 2004). 模拟氮沉降对三种南亚热带树苗生长和光合作用的影响. 生态学报, 24, 876-882.]
DOI URL |
|
| [24] |
Li XF, Zheng XB, Han SJ, Zheng JQ, Li TH ( 2010). Effects of nitrogen additions on nitrogen resorption and use efficiencies and foliar litterfall of six tree species in a mixed birch and poplar forest, northeastern China. Canadian Journal of Forest Research, 40, 2256-2261.
DOI URL |
| [25] |
Lin BP, He ZM, Lin SZ, Hu HT, Qiu LJ, Liu ZM ( 2017). Needles macronutrient concentrations and retranslocation characteristics in Chinese fir plantation of different ages. Journal of Forest and Environment, 37, 34-39.
DOI URL |
|
[ 林宝平, 何宗明, 林思祖, 胡欢甜, 邱岭军, 刘卓明 ( 2017). 不同林龄杉木针叶大量元素转移特征. 森林与环境学报, 37, 34-39.]
DOI URL |
|
| [26] |
Liu WF, Fan HB, Zhang ZW, Yang YL, Wang QQ, Xu L ( 2008). Foliar nutrient contents of Chinese fir in response to simulated nitrogen deposition. Chinese Journal of Applied & Environment Biology, 14, 319-323.
DOI URL |
|
[ 刘文飞, 樊后保, 张子文, 杨跃霖, 王启其, 徐雷 ( 2008). 杉木人工林针叶养分含量对模拟氮沉降增加的响应. 应用与环境生物学报, 14, 319-323.]
DOI URL |
|
| [27] |
Liu XJ, Zhang Y, Han WX, Tang AH, Shen LJ, Cui ZL, Vitousek P, Erisman JW, Goulding K, Christie P, Andreas FM, Zhang FS ( 2013). Enhanced nitrogen deposition over China. Nature, 494, 459-462.
DOI URL PMID |
| [28] |
Lu JY, Duan BH, Yang M, Yang H, Yang HM ( 2018). Research process in nitrogen and phosphorus resorption from senesced leaves and the influence of ontogenetic and environmental factors. Acta Prataculturae Sinca, 27, 178-188.
DOI URL |
|
[ 陆姣云, 段兵红, 杨梅, 杨晗, 杨慧敏 ( 2018). 植物叶片氮磷养分重吸收规律及其调控机制研究进展. 草业学报, 27, 178-188.]
DOI URL |
|
| [29] |
Matson P, Lohse KA, Hall SJ ( 2002). The globalization of nitrogen deposition: Consequences for terrestrial ecosystems. AMBIO, 31, 113-119.
DOI URL PMID |
| [30] |
Mayor JR, Wright SJ, Turner BL ( 2014). Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest. Journal of Ecology, 102, 36-44.
DOI URL |
| [31] |
Ning QR, Li SZ, Jiang LC, Zhao Y, Liu R, Zhang XY ( 2016). Foliar nutrient content and resorption efficiency of Pinus massoniana in the subtropical red soil erosion region. Acta Ecologica Sinica, 36, 3510-3517.
DOI URL |
|
[ 宁秋蕊, 李守中, 姜良超, 赵颖, 刘溶, 张欣影 ( 2016). 亚热带红壤侵蚀区马尾松针叶养分含量及再吸收特征. 生态学报, 36, 3510-3517.]
DOI URL |
|
| [32] |
Reed SC, Townsend AR, Davidson EA, Cleveland CC ( 2012). Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytologist, 196, 173-180.
DOI URL PMID |
| [33] |
Reich PB, Oleksyn J, Wright IJ ( 2009). Leaf phosphorus influences the photosynthesis-nitrogen relation: A cross-biome analysis of 314 species. Oecologia, 160, 207-212.
DOI URL PMID |
| [34] | Shen FF, Wu JP, Fan HB, Guo XM, Lei XM, Wo QD ( 2018). Litterfall ecological stoichiometry and soil available nutrients under long-term nitrogen deposition in a Chinese fir plantation. Acta Ecologica Sinica, 38, 7477-7487. |
| [ 沈芳芳, 吴建平, 樊后保, 郭晓敏, 雷学明, 沃奇东 ( 2018). 杉木人工林凋落物生态化学计量与土壤有效养分对长期模拟氮沉降的响应. 生态学报, 38, 7477-7487.] | |
| [35] |
Sohrt J, Herschbach C, Weiler M ( 2018). Foliar P- but not N resorption efficiency depends on the P-concentration and the N:P ratio in trees of temperate forests. Trees, 32, 1443-1445.
DOI URL |
| [36] |
van Heerwaarden LM, Toet S, Aerts R ( 2003). Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization. Journal of Ecology, 91, 1060-1070.
DOI URL |
| [37] |
Vergutz L, Manzoni S, Porporato A, Novais RF, Jackson RB ( 2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecological Monographs, 82, 205-220.
DOI URL |
| [38] | Wang B, Huang G, Ma J, Li Y ( 2016). Responses of nutrients resorption of five desert ephemeral plants to water and nitrogen additions. Journal of Desert Research, 36, 415-422. |
| [ 王斌, 黄刚, 马健, 李彦 ( 2016). 5种荒漠短命植物养分再吸收对水氮添加的响应. 中国沙漠, 36, 415-422.] | |
| [39] |
Wang M, Murphy MT, Moore TR ( 2014). Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog. Oecologia, 174, 365-377.
DOI URL PMID |
| [40] |
Wang R, Goll D, Balkanski Y, Hauglustaine D, Boucher O, Ciais P, Janssens I, Penuelas J, Guenet B, Sardans J, Bopp L, Vuichard N, Zhou F, Li BG, Piao SL, Peng SS, Huang Y, Tao S ( 2017). Global forest carbon uptake due to nitrogen and phosphorus deposition from 1850 to 2100. Global Change Biology, 23, 4854-4872.
DOI URL PMID |
| [41] |
Wright IJ, Leishman MR, Read C, Westoby M ( 2006). Gradients of light availability and leaf traits with leaf age and canopy position in 28 Australian shrubs and trees. Functional Plant Biology, 33, 407-419.
DOI URL |
| [42] |
Wright I J, Westoby M ( 2003). Nutrient concentration, resorption and lifespan: Leaf traits of Australian sclerophyll species. Functional Ecology, 17, 10-19.
DOI URL |
| [43] |
Wu JP, Duan HL, Liu WF, Wei XH, Liao YC, Fan HB ( 2017). Individual size but not additional nitrogen regulates tree carbon sequestration in a subtropical forest. Scientific Reports, 7, 46293. DOI: 10.1038/srep46293.
DOI URL PMID |
| [44] |
Wu PF, Ma XQ ( 2009). Research advances in the mechanisms of high nutrient use efficiency in plants. Acta Ecologica Sinica, 29, 427-437.
DOI URL |
|
[ 吴鹏飞, 马祥庆 ( 2009). 植物养分高效利用机制研究进展. 生态学报, 29, 427-437.]
DOI URL |
|
| [45] |
Xiao J ( 2005). Formation and damage of atmospheric wet deposition of nitrogen in Zhangzhou. Energy and Enviroment, 2, 59-61.
DOI URL |
|
[ 肖健 ( 2005). 漳州市氮湿沉降量异常的形成及危害. 能源与环境, 2, 59-61.]
DOI URL |
|
| [46] |
Yan ZB, Kim NY, Han XW, Guo YL, Han TS, Du EZ, Fang JY ( 2014). Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant and Soil, 388, 147-155.
DOI URL |
| [47] |
Yuan ZY, Chen HYH ( 2009). Global trends in senesced-leaf nitrogen and phosphorus. Global Ecology and Biogeogrphy, 18, 532-542.
DOI URL |
| [48] |
Yuan ZY, Li LH, Han XG, Huang JH, Jiang GM, Wan SQ, Zhang WH, Chen QS ( 2005). Nitrogen resorption from senescing leaves in 28 plant species in a semi-arid region of northern China. Journal of Arid Environments, 63, 191-202.
DOI URL |
| [49] |
Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Pe?uelas J, Richiter A, Sardans J, Wanek W ( 2015). The application of ecological stoichiometry to plant-microbial- soil organic matter transformations. Ecological Monographs, 85, 133-155.
DOI URL |
| [50] |
Zhang JH, Tang YZ, Luo YK, Chi XL, Chen YH, Fang JY, Shen HH ( 2014). Resorption efficiency of leaf nutrients in woody plants on Mt. Dongling of Beijing, North China. Journal of Plant Ecology, 8, 530-538.
DOI URL |
| [51] | Zhang LY, Zhang DP ( 2003). The phloem unloading pathway and mechanism of photoassimilates. Plant Physiology Communications, 39, 399-403. |
| [ 张凌云, 张大鹏 ( 2003). 光合同化物韧皮部卸载途径和机制. 植物生理学报, 39, 399-403.] | |
| [52] |
Zhang QF, Xie JS, Chen NS, Chen T, Lü MK, Zhang H, Yang YS ( 2017). Effects of ecological restoration on stoichiometric characteristics and nutrient resorption efficiency of Pinus massoniana foliage. Acta Ecologica Sinica, 37, 267-276.
DOI URL |
|
[ 张秋芳, 谢锦升, 陈奶寿, 陈坦, 吕茂奎, 张浩, 杨玉盛 ( 2017). 生态恢复对马尾松叶片化学计量及氮磷转移的影响. 生态学报, 37, 267-276.]
DOI URL |
|
| [53] |
Zhao Q, Liu XY, Hu YL, Zeng DH ( 2010). Effects of nitrogen addition on nutrient allocation and nutrient resorption efficiency in Larix gmelinii. Scientia Silvae Sinicae, 46(5), 14-19.
DOI URL |
|
[ 赵琼, 刘兴宇, 胡亚林, 曾德慧 ( 2010). 氮添加对兴安落叶松养分分配和再吸收效率的影响. 林业科学, 46(5), 14-19.]
DOI URL |
| [1] | 张文瑾 佘维维 秦树高 乔艳桂 张宇清. 氮和水分添加对黑沙蒿群落优势植物叶片氮磷化学计量特征的影响[J]. 植物生态学报, 2024, 48(5): 590-600. |
| [2] | 韩路, 冯宇, 李沅楷, 王雨晴, 王海珍. 地下水埋深对灰胡杨叶片与土壤养分生态化学计量特征及其内稳态的影响[J]. 植物生态学报, 2024, 48(1): 92-102. |
| [3] | 李兆光, 文高, 和桂青, 徐天才, 和琼姬, 侯志江, 李燕, 薛润光. 滇西北藜麦氮磷钾生态化学计量特征的物候期动态[J]. 植物生态学报, 2023, 47(5): 724-732. |
| [4] | 刘婧, 缑倩倩, 王国华, 赵峰侠. 晋西北丘陵风沙区柠条锦鸡儿叶片与土壤生态化学计量特征[J]. 植物生态学报, 2023, 47(4): 546-558. |
| [5] | 林少颖, 曾瑜, 杨文文, 陈斌, 阮敏敏, 尹晓雷, 阳祥, 王维奇. 添加秸秆及其生物炭对茉莉植株与土壤碳氮磷生态化学计量特征的影响[J]. 植物生态学报, 2023, 47(4): 530-545. |
| [6] | 尹晓雷, 刘旭阳, 金强, 李先德, 林少颖, 阳祥, 王维奇, 张永勋. 不同管理模式对茶树碳氮磷含量及其生态化学计量比的影响[J]. 植物生态学报, 2021, 45(7): 749-759. |
| [7] | 朱湾湾, 王攀, 许艺馨, 李春环, 余海龙, 黄菊莹. 降水量变化与氮添加下荒漠草原土壤酶活性及其影响因素[J]. 植物生态学报, 2021, 45(3): 309-320. |
| [8] | 胡琪娟, 盛茂银, 殷婕, 白义鑫. 西南喀斯特石漠化环境适生植物构树细根、根际土壤化学计量特征[J]. 植物生态学报, 2020, 44(9): 962-972. |
| [9] | 解梦怡, 冯秀秀, 马寰菲, 胡汗, 王洁莹, 郭垚鑫, 任成杰, 王俊, 赵发珠. 秦岭锐齿栎林土壤酶活性与化学计量比变化特征及其影响因素[J]. 植物生态学报, 2020, 44(8): 885-894. |
| [10] | 刘珊杉, 周文君, 况露辉, 刘占锋, 宋清海, 刘运通, 张一平, 鲁志云, 沙丽清. 亚热带常绿阔叶林土壤胞外酶活性对碳输入变化及增温的响应[J]. 植物生态学报, 2020, 44(12): 1262-1272. |
| [11] | 熊星烁, 蔡宏宇, 李耀琪, 马文红, 牛克昌, 陈迪马, 刘娜娜, 苏香燕, 景鹤影, 冯晓娟, 曾辉, 王志恒. 内蒙古典型草原植物叶片碳氮磷化学计量特征的季节动态[J]. 植物生态学报, 2020, 44(11): 1138-1153. |
| [12] | 李军军, 李萌茹, 齐兴娥, 王立龙, 徐世健. 芨芨草叶片养分特征对氮磷不同添加水平的响应[J]. 植物生态学报, 2020, 44(10): 1050-1058. |
| [13] | 贾丙瑞. 凋落物分解及其影响机制[J]. 植物生态学报, 2019, 43(8): 648-657. |
| [14] | 王攀, 朱湾湾, 牛玉斌, 樊瑾, 余海龙, 赖江山, 黄菊莹. 氮添加对荒漠草原植物群落组成与微生物生物量生态化学计量特征的影响[J]. 植物生态学报, 2019, 43(5): 427-436. |
| [15] | 杨文高, 字洪标, 陈科宇, 阿的鲁骥, 胡雷, 王鑫, 王根绪, 王长庭. 青海森林生态系统中灌木层和土壤生态化学计量特征[J]. 植物生态学报, 2019, 43(4): 352-364. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||
Copyright © 2022 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19